Суммация сокращений и тетанус
Если в эксперименте на отдельное мышечное волокно или на всю мышцу действуют два быстро следующих друг за другом сильных одиночных раздражения, то возникающее сокращение будет иметь большую амплитуду, чем максимальное сокращение при одиночном раздражении. Сократительные эффекты, вызванные первым и вторым раздражениями, как бы складываются. Это явление называется суммацией сокращений (рис. 29). Оно наблюдается как при прямом, так и при непрямом раздражении мышцы.
Для возникновения суммации необходимо, чтобы интервал между раздражением имел определенную длительность: он должен быть длиннее рефрактерного периода, иначе на второе раздражение не будет ответа, и короче всей длительности сократительного ответа, чтобы второе раздражение подействовало на мышцу раньше, чем она успеет расслабиться после первого раздражения. При этом возможны два варианта.
Если второе раздражение поступает, когда мышца уже начала расслабляться, то на миографической кривой вершина этого сокращения будет отделена от вершины первого западенисм (рис. 29, Ж — Г). Если же второе раздражение действует, когда первое еще не дошло до своей вершины, то второе сокращение полностью сливается с первым, образуя единую суммированную вершину (рис. 29, А — В).


Как при полной, так и при неполной суммации сокращений потенциалы действия не суммируются.
Тетанус мышцы. Если на отдельное мышечное волокно или на всю мышцу действуют ритмические раздражения с такой частотой, что их эффекты суммируются, наступает сильное и длительное сокращение мышцы, называемое тетаническим сокращением, или тетанусом. Амплитуда его может быть в несколько раз больше величины максимального одиночного сокращения.
После прекращения тетанического раздражения волокна полностью расслабляются, их исходная длина восстанавливается лишь по истечении некоторого времени. Это явление называется цоелстетанической, или остаточной, контрактурой.
Чем быстрее сокращаются и расслабляются волокна мышцы, тем чаще должны быть раздражения, чтобы вызвать тетанус.
Физиология человека. Общая. Спортивная. Возрастная (15 стр.)

Рис. 11. Одиночное сокращение, зубчатый и сплошной тетанус камбаловидной мышцы человека
(по: Н.В. Зимкин и др., 1984); верхняя кривая – сокращение мышцы, нижняя – отметка раздражения мышцы, справа указана частота раздражения
Если интервалы между нервными импульсами короче, чем длительность одиночного сокращения, то возникает явление суперпозиции – наложение механических эффектов мышечного волокна друг на друга и наблюдается сложная форма сокращения – тетанус. Различают две формы тетануса – зубчатый тетанус, возникающий при более редком раздражении, когда происходит попадание каждого следующего нервного импульса в фазу расслабления отдельных одиночных сокращений, и сплошной, или гладкий, тетанус, возникающий при более частом раздражении, когда каждый следующий импульс попадает в фазу сокращения (рис. 11). Таким образом (в некоторых границах) между частотой импульсов возбуждения и амплитудой сокращения волокон ДЕ существует определенное соотношение: при небольшой частоте (например, 5–8 имп. в 1 с) возникают одиночные сокращения, при увеличении частоты (15–20 имп. в 1 с) – зубчатый тетанус, при дальнейшем нарастании частоты (25–60 имп. в 1 с) – гладкий тетанус. Одиночное сокращение – более слабое и менее утомительное, чем тетаническое. Зато тетанус обеспечивает в несколько раз более мощное, хотя и кратковременное сокращение мышечного волокна.
Сокращение целой мышцы зависит от формы сокращения отдельных ДЕ и их координации во времени. При обеспечении длительной, но не очень интенсивной работы, отдельные ДЕ сокращаются попеременно (рис. 12), поддерживая общее напряжение мышцы на заданном уровне (например, при беге на длинные и сверхдлинные дистанции). При этом отдельные ДЕ могут развивать как одиночные, так и тетанические сокращения, что зависит от частоты нервных импульсов. Утомление в этом случае развивается медленно, так как, работая по очереди, ДЕ в промежутках между активацией успевают восстанавливаться. Однако для мощного кратковременного усилия (например, поднятия штанги) требуется синхронизация активности отдельных ДЕ, т. е. одновременное возбуждение практически всех ДЕ, что, в свою очередь, требует одновременной активации соответствующих нервных центров и достигается в результате длительной тренировки. При этом осуществляется мощное и весьма утомительное тетаническое сокращение.

Рис. 12. Различные режимы работы двигательных единиц (ДЕ)
Амплитуда сокращения одиночного волокна не зависит от силы надпорогового раздражения (закон «Все или ничего»), В отличие от этого, при нарастании силы надпорогового раздражения сокращение целой мышцы постепенно растет до максимальной амплитуды.
Работа мышцы с небольшой нагрузкой сопровождается редкой частотой нервных импульсов и вовлечением небольшого числа ДЕ. В этих условиях, накладывая отводящие электроды на кожу над мышцей и используя усилительную аппаратуру, можно на экране осциллографа или с применением чернильной записи на бумаге зарегистрировать одиночные потенциалы действия отдельных ДЕ. В случае же значительных напряжений потенциалы действия многих ДЕ. алгебраически суммируются, возникает сложная интегрированная кривая записи электрической активности целой мышцы – электромиограмма (ЭМГ).
Форма ЭМГ отражает характер работы мышцы: при статических усилиях она имеет непрерывный вид, а при динамической работе – вид отдельных пачек импульсов, приуроченных в основном к начальному моменту сокращения мышцы и разделенных периодами «электрического молчания». Особенно хорошо ритмичность появления подобных пачек наблюдается у спортсменов при циклической работе (рис. 13). У маленьких детей и неадаптированных к такой работе лиц четких периодов отдыха не наблюдается, что отражает недостаточное расслабление мышечных волокон работающей мышцы.
Чем больше внешняя нагрузка и сила сокращения мышцы, тем выше амплитуда ее ЭМГ. Это связано с увеличением частоты нервных импульсов, вовлечением большего числа ДЕ в мышце и синхронизацией их активности. Современная многоканальная аппаратура позволяет производить одновременную регистрацию ЭМГ многих мышц на разных каналах. При выполнении спортсменом сложных движений можно видеть на полученных ЭМГ кривых не только характер активности отдельных мышц, но и оценить моменты и порядок их включения или выключения в различные фазы двигательных актов. Записи ЭМГ, полученные в естественных условиях двигательной деятельности, можно передавать к регистрирующей аппаратуре по телефону или радиотелеметрически. Анализ частоты, амплитуды и формы ЭМГ (например, с помощью специальных компьютерных программ) позволяет получить важную информацию об особенностях техники выполняемого спортивного упражнения и степени ее освоения обследуемым спортсменом.

Рис. 13. Электромиограмма мышц-антагонистов при циклической работе
По мере развития утомления при той же величине мышечного усилия амплитуда ЭМГ нарастает. Это связано с тем, что снижение сократительной способности утомленных ДЕ компенсируется нервными центрами вовлечением в работу дополнительных ДЕ, т. е. путем увеличения количества активных мышечных волокон. Кроме того, усиливается синхронизация активности ДЕ, что также повышает амплитуду суммарной ЭМГ.
5.4. монофункциональные основы мышечной силы
Движение является результатом взаимодействия внутренних и внешних сил, развиваемых в опорно-двигательном аппарате, – активных (возникающих при сокращении или напряжении мышцы во время ее возбуждения) и пассивных (упругое напряжение при растяжении мышцы, сопротивление мышцы и ее сухожилия).
Сила мышцы зависит от ряда морфологических и физиологических факторов: количества и свойств мышечных волокон в мышце, исходной длины мышцы, характера нервных импульсов, механических условий действия мышцы на кости скелета.
Сила мышцы является суммой силы отдельных ее мышечных волокон. Подсчитано, что 1 одиночное мышечное волокно икроножной мышцы развивает напряжение 100–200 мг, 1 ДЕ икроножной мышцы человека содержит около 2000 мышечных волокон и развивает напряжение 200–400 г, 1 икроножная мышца содержит около 1000 ДЕ и развивает напряжение 200–400 кг.
Большое значение имеет анатомическое строение мышцы. В параллельно-волокнистых и веретенообразных мышцах (камбаловидная мышца и др.) сила мышц тем больше, чем больше ее анатомический поперечник, т. е. площадь поперечного сечения целой мышцы. В перистых мышцах (двуглавая мышца и др.) физиологический поперечник, т. е. площадь поперечного сечения всех мышечных волокон, гораздо больше, чем ее анатомический поперечник. В такой мышце упаковано значительно больше мышечных волокон и соответственно больше ее сила.
На силу сокращения мышцы влияет ее исходная длина, так как от нее зависит возможное количество поперечных мостиков между актином и миозином. Предполагают, что в каждом цикле присоединения-отсоединения поперечных мостиков расходуется энергия 1 молекулы АТФ на 1 поперечный мостик. Следовательно, чем больше образуется в мышечном волокне актино-миозиновых мостиков, тем выше скорость расщепления АТФ, больше тяга сократительных белков и соответственно больше развиваемая мышцей сила.
Наибольшее количество актино-миозиновых контактов образуется при небольшом растяжении мышцы до некоторой оптимальной длины. При значительном растяжении саркомера нити актина далеко расходятся в стороны и практически не контактируют с расположенным в средней части саркомера миозином. В случае же резкого уменьшения длины саркомера нити актина в центре перекрывают друг друга, препятствуя контактам с миозином и также уменьшая число образуемых мостиков. В связи с этими особенностями взаимодействия сократительных белков наибольшая сила мышцы проявляется при некотором ее предварительном растяжении.
Одной из важнейших характеристик скелетных мышц, влияющих на силу сокращения, является состав (композиция) мышечных волокон. Различают три типа мышечных волокон – медленные неутомляемые (I типа), быстрые неутомляемые или промежуточные (II-а типа) и быстрые утомляемые (II – б типа).
Медленные волокна (I типа), их обозначают также SO – Slow Oxydative (англ. – «медленные окислительные»), – это выносливые (неутомляемые) и легко возбудимые волокна, с богатым кровоснабжением, большим количеством митохондрий, запасов миоглобина и с использованием окислительных процессов энергообразования (аэробные). Их у человека в среднем 50 %. Они легко включаются в работу при малейших напряжениях мышц, очень выносливы, но не обладают достаточной силой. Чаще всего они используются при поддержании ненагрузочной статической работы, например при сохранении позы.
Одиночное сокращение, суммация, тетанус




![]()
![]()
Биомеханика мышечных сокращений.
При нанесении на двигательный нерв или мышцу одиночного порогового или сверхпорогового раздражения, возникает одиночное сокращение. При его графической регистрации, на полученной кривой можно выделить три последовательных периода:
1.Латентный период. Это время от момента нанесения раздражения до начала сокращения. Его длительность около 1-2 мсек. Во время латентного периода генерируется и распространяется ПД, происходит высвобождения кальция из СР, взаимодействие актина с миозином и т.д.
2.Период укорочения. В зависимости от типа мышцы (быстрая или медленная) его продолжительность от 10 до 100 мсек.,
3.Период расслабления. Его длительность несколько больше, чем укорочения. Рис.
В режиме одиночного сокращения мышца способна работать длительное время без утомления, но его сила незначительна. Поэтому в организме такие сокращения встречаются редко, например так могут сокращаться быстрые глазодвигательные мышцы. Чаще одиночные сокращения суммируются.
Суммация это сложение 2-х последовательных сокращений при нанесении на нее 2-х пороговых или сверхпороговых раздражений, интервал между которыми меньше длительности одиночного сокращения, но больше продолжительности рефрактерного периода. Различают 2 вида суммации: полную и неполную суммацию. Неполная суммация возникает в том случае, если повторное раздражение наносится на мышцу, когда он уже начала расслабляться. Полная возникает тогда, когда повторное раздражение действует на мышцу до начала периода расслабления, т.е. в конце периода укорочения.(рис 1,2). Амплитуда сокращения при полной суммации выше, чем неполной. Если интервал между двумя раздражениями еще больше уменьшить. Например, нанести второе в середине периода укорочения, то суммации не будет, потому что мышца находится в состоянии рефрактерности.
Тетанус— это длительное сокращение мышцы, возникающее в результате суммации нескольких одиночных сокращений, развивающихся при нанесении на нее ряда последовательных раздражений. Различают 2 формы тетануса: зубчатый и гладкий. Зубчатый тетанус наблюдается в том случае, если каждое последующее раздражение действует на мышцу, когда она уже начала расслабляться. Т.е. наблюдается неполная суммация (рис). Гладкий тетанус возникает тогда, когда каждое последующее раздражение наносится в конце периода укорочения. Т.е. имеет место полная суммация отдельных сокращений и (рис). Амплитуда гладкого тетануса больше, чем зубчатого. В норме мышцы человека сокращаются в режиме гладкого тетануса. Зубчатый возникает при патологии, например тремор рук при алкогольной интоксикации и болезни Паркинсона.
Тетаническое сокращение мышцы
![]()
![]()
При воздействии на мышцу ритмических раздражений высокой частоты наступает сильное и длительное сокращение мышцы, которое называется тетаническим сокращением или тетанусом. Этот термин впервые применил Э. Вебер в 1821 году.
Тетанус может быть зубчатым (при частоте раздражений 20-40 Гц) или сплошным, гладким (при частоте 50 Гц и выше). Амплитуда тетанического сокращения в 2–4 раза выше амплитуды одиночного сокращения при той же силе раздражения.
I 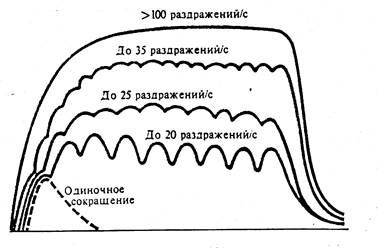
II 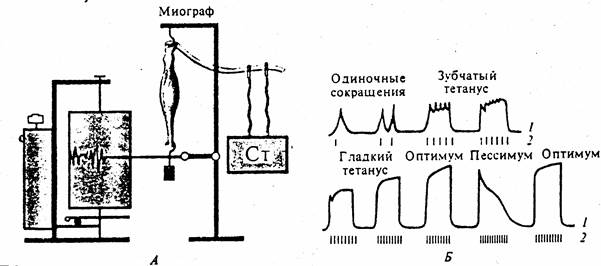
Примечание. Явления пессимума и парабиоза возможны
в условиях эксперимента.
Рис. 81. Формирование тетануса в зависимости от частоты раздражения.
Оптимум и пессимум (по Н.Введенскому). А- схема регистрации; Б- кривые мышечных сокращений (1) при различной частоте раздражений (2).
Гладкий тетанус возникает тогда, когда очередной импульс раздражения действует на мышцу до начала фазы расслабления. При очень большой частоте раздражений каждое очередное раздражение будет попадать на фазу абсолютной рефрактерности и мышца вообще не будет сокращаться. Высота мышечного сокращения при тетанусе зависит от ритма раздражения, а также от возбудимости и лабильности, которые изменяются в процессе сокращения мышцы. Тетанус наиболее высокий при оптимальном ритме, когда каждый последующий импульс действует на мышцу в фазу экзальтации, вызванной предыдущим импульсом. В этом случае создаются наилучшие условия (оптимум силы и частоты раздражения, оптимум ритма) для работы мышцы.
При тетанических сокращениях мышечные волокна утомляются больше, чем при одиночных сокращениях. Поэтому даже в пределах одной мышцы происходит периодическая смена частоты импульсации (вплоть до полного исчезновения) в разных двигательных единицах.
Импульсы с мотонейронов в условиях покоя участвуют в поддержании мышечного тонуса.
Под тонусом понимают состояние естественного постоянного напряжения мышц при невысоких энергетических затратах. В поддержании тонуса участвуют проприорецепторы мышц (мышечные веретена) и центральная нервная система.
Осуществление тонуса скелетных мышц обусловлено функцией медленных двигательных единиц красных волокон мышц. Тонус скелетных мышц связан с поступлением редких нервных импульсов к мышце, в результате чего мышечные волокна возбуждаются не одновременно, а попеременно. У домашних животных существуют специализированные рефлекторные дуги, одни из которых обеспечивают тетанические сокращения, а другие мышечный тонус. Тонус скелетных мышц играет важную роль в поддержании определенного положения тела в пространстве и деятельности двигательного аппарата.
Нормальная физиология (7 стр.)

Если каждый последующий стимул поступает к мышце в тот период, когда она находится в фазе укорочения, то возникает гладкий тетанус, а если в фазу расслабления – зубчатый тетанус.
Амплитуда тетанического сокращения превышает амплитуду одиночного мышечного сокращения. Н. Е. Введенский объяснил это явление фазными изменениями возбудимости мышцы, введя понятие об оптимуме и пессимуме частоты раздражения. Оптимум – такая частота раздражения, при которой каждое последующее раздражение наносится в фазу повышенной возбудимости. Тетанус при этом будет максимальным по амплитуде. Пессимум – такая частота раздражения, при которой каждое последующее раздражение наносится в фазу пониженной возбудимости. Амплитуда тетануса при этом будет минимальной.
Различают несколько видов мышечных сокращений: изотонический, изометрический и смешанный. При изотоническом сокращении мышцы происходит изменение ее длины, а напряжение остается постоянным. Такое сокращение происходит в том случае, если отсутствует сопротивление изменению ее длины. К изотоническому типу сокращений относятся сокращения мышц языка. При изометрическом сокращении длина мышечных волокон остается постоянной, а их напряжение возрастает. Такое сокращение мышцы возникает при попытке поднять чрезмерно большой груз. В естественных условиях сокращения мышц никогда не бывают чисто изотоническими или изометрическими, они имеют смешанный характер, т. е. происходит изменение и длины, и напряжения мышцы.
Механизм мышечного сокращения
Мышцы состоят из мышечных волокон, а те – из множества тонких нитей – миофибрилл, расположенных продольно. Каждая миофибрилла состоит из нитей сократительных белков актина и миозина. Перегородки, называемые Z-пластинами, разделяют миофибриллы на участки – саркомеры. В саркомере чередуются поперечные светлые и темные полосы. Поперечная исчерченность миофибрилл обусловлена определенным расположением нитей актина и миозина. В центральной части каждого саркомера расположены толстые нити миозина. На обоих концах саркомера находятся тонкие нити актина, прикрепленные к Z-пластинам. Нити миозина выглядят в световом микроскопе как светлая полоска (Н-зона) в темном диске, который содержит нити миозина и актина и называется анизотропным, или А-диском. По обе стороны от А-диска находятся участки, которые содержат только тонкие нити актина и кажутся светлыми, они называются изотропными, или I-дисками. По их середине проходит темная линия – Z-мембрана. Благодаря такому периодическому чередованию светлых и темных дисков сердечная и скелетная мышцы выглядят поперечно-полосатыми (рис.7).
В состоянии покоя концы толстых и тонких нитей лишь незначительно перекрываются на уровне А-диска. При сокращении тонкие актиновые нити скользят вдоль толстых миозиновых нитей, двигаясь между ними к середине саркомера. Сами актиновые и миозиновые нити своей длины не изменяют. Миозиновые нити имеют поперечные мостики (выступы) с головками, которые отходят от нити биполярно. Актиновая нить состоит из двух закрученных одна вокруг другой цепочек молекул актина. На нитях актина расположены молекулы тропонина, а в желобках между двумя актина лежат нити тропомиозина. Молекулы тропомиозина в покое располагаются так, что предотвращают прикрепление поперечных мостиков миозина к актиновым нитям.

Во многих местах участки поверхностной мембраны мышечной клетки углубляются в виде трубочек внутрь волокна, перпендикулярно его продольной оси, образуя систему поперечных трубочек (Т-систему). Параллельно миофибриллам и перпендикулярно поперечным трубочкам расположена система продольных трубочек (альфа-система). Пузырьки на концах этих трубочек, в которых сосредоточено основное количество внутриклеточного кальция, подходят очень близко к поперечным трубочкам, образуя совместно с ними так называемые триады. В состоянии покоя миозиновый мостик заряжен энергией (миозин фосфорилирован), но он не может соединиться с нитью актина, так как между ними находится система из нитей тропомиозина и молекул тропонина. При возбуждении ПД распространяется по мембранам Тсистемы внутрь клетки и вызывает высвобождение ионов кальция из альфа-системы. С появлением ионов кальция в присутствии АТФ происходит изменение пространственного положения тропонина – нить тропомиозина сдвигается и открываются участки актина, присоединяющие миозиновые головки. Соединение головки фосфорилированного миозина с актином приводит к изменению положения мостика (его «сгибанию»), в результате нити актина перемещаются на 1 мм к середине саркомера. Затем происходит отсоединение мостика от актина. Ритмические прикрепления и отсоединения головок миозина тянут актиновую нить к середине саркомера.
При отсутствии повторного возбуждения ионы кальция закачиваются кальциевым насосом из межфибриллярного пространства в систему саркоплазматического ретикулума. Это приводит к снижению концентрации ионов кальция и отсоединению его от тропонина. Вследствие чего тропомиозин возвращается на прежнее место и снова блокирует активные центры актина. Затем происходит фосфорилирование миозина за счет АТФ, что также способствует временному разобщению нитей. Расслабление мышцы после ее сокращения происходит пассивно – актиновые и миозиновые нити легко скользят в обратном направлении под влиянием сил упругости мышечных волокон, а также сокращения мышц-антагонистов.
Гладкие мышцы
Эти мышцы образуют мышечные слои стенок желудка, кишечника, мочеточников, бронхов, кровеносных сосудов и других внутренних органов. Они построены из веретенообразных одноядерных мышечных клеток. Гладкие мышцы разделяются на две основные группы: мультиунитарные и унитарные. Мультиунитарные мышцы функционируют независимо друг от друга, и каждое волокно может иннервироваться отдельным нервным окончанием. Такие волокна обнаружены в ресничной мышце глаза, мигательной перепонке и мышечных слоях некоторых крупных сосудов, к ним относятся мышцы, поднимающие волосы. У унитарных мышц волокна настолько тесно переплетены, что их мембраны могуг сливаться, образуя электрические контакты (нексусы). При раздражении одного волокна за счет этих контактов ПД быстро распространяются на соседние волокна. Поэтому, несмотря на то, что двигательные нервные окончания расположены на небольшом числе мышечных волокон, в реакцию вовлекается вся мышца. Такие мышцы имеются в большинстве органов: пищеварительном тракте, матке, в мочеточниках.
Особенностью гладких мышц является их способность осуществлять медленные и длительные тонические сокращения. Медленные, ритмические сокращения гладких мышц желудка, кишечника, мочеточников и других органов обеспечивают перемещение содержимого этих органов. Длительные тонические сокращения гладких мышц обеспечивают функционирование сфинктеров полых органов, которые препятствуют выходу их содержимого.
Гладкие мышцы стенок кровеносных сосудов, особенно артерий и артериол, также находятся в состоянии постоянного тонического сокращения. Изменение тонуса мышц стенок артериальных сосудов влияет на величину их просвета и, следовательно, на уровень кровяного давления и кровоснабжения органов. Важным свойством гладких мышц является их пластичность, т. е. способность сохранять приданную им при растяжении длину. Скелетная мышца в норме почти не обладает пластичностью. При удалении растягивающего груза скелетная мышца быстро укорачивается, а гладкая остается растянутой. Высокая пластичность гладких мышц имеет большое значение для нормального функционирования полых органов. Например, пластичность мышц мочевого пузыря по мере его наполнения предотвращает избыточное повышение давления.
Сильное и резкое растяжение гладких мышц вызывает их сокращение, что обусловлено нарастающей при растяжении деполяризацией клеток, которая обеспечивает автоматию гладкой мышцы. Такое сокращение играет важную роль в авторегуляции тонуса кровеносных сосудов, а также способствует непроизвольному опорожнению переполненного мочевого пузыря в тех случаях, когда нервная регуляция отсутствует в результате повреждения спинного мозга.
В гладких мышцах тетаническое сокращение возникает при низкой частоте стимуляции. В отличие от скелетных, гладкие мышцы способны развивать спонтанные тетанообразные сокращения в условиях денервации и даже после блокады интрамуральных ганглиев. Такие сокращения возникают вследствие активности клеток, обладающих автоматией (пейсмекерных клеток), которые отличаются по электрофизиологическим свойствам от других мышечных клеток. В них появляются пейсмекерные потенциалы, деполяризующие мембрану до критического уровня, что вызывает возникновение потенциала действия.
Особенностью гладких мышц является их высокая чувствительность к медиаторам, которые оказывают на спонтанную активность пейсмекеров модулирующие влияния. При нанесении ацетилхолина на препарат мышцы толстой кишки частота ПД возрастает. Вызываемые ими сокращения сливаются, образуется почти гладкий тетанус. Чем выше частота ПД, тем сильнее сокращение. Норадреналин, напротив, гиперполяризует мембрану, снижая частоту ПД и величину тетануса.
Возбуждение гладкомышечных клеток вызывает повышение концентрации кальция в саркоплазме, что активирует сократительные структуры. Так же как сердечная и скелетная мышцы, гладкая мышца расслабляется при снижении концентрации ионов кальция. Расслабление гладких мышц происходит медленнее, так как удаление ионов кальция замедлено.