Почему амплитуда тетануса скелетной мышцы больше чем амплитуда одиночного сокращения

Известно, что во время длительного сокращения мышцы, регистрируемая механическая сила градуально снижается. Этот феномен, рассматриваемый как «утомление», один из наиболее интригующих явлений, ассоциируемых с сократительной активностью мышцы. Процесс утомления скелетных мышц исследовали в разных экспериментальных условиях и в большинстве случаев определяли, как неспособность поддерживать требуемую или рассчитанную силу «an inability to maintain the required or expected force» [19]), или как точка нарушения «the failure point» [31]). Несмотря на многочисленные исследования, этиология мышечного утомления остается неизвестной [22], а тем более в условиях механической разгрузки мышечного аппарата.
В качестве показателя мышечного утомления широко используют изменения поверхностной электромиограммы (ЭМГ) 26. Однако связь между ЭМГ и утомлением не только нелинейная, но часто изменения в ЭМГ предшествуют утомлению мышцы [27] и интерпретация ЭМГ сигнала затруднена сложностью его происхождения. Параметры ЭМГ во время произвольной активности мышцы являются результатом суммации и «выпадением» отдельных потенциалов действия (ПД) двигательных единиц из числа всех активных двигательных единиц [16]. Известно, что характеристика ЭМГ зависит от паттерн частоты импульсации двигательной единицы и конфигурации ПД [28]. В той связи объективным, легко измеряемым, физиологическим показателем оценки утомления является активность мышцы при электрической стимуляции постоянной частотой. В последнем случае параметры механического ответа не зависят от центрального драйва [14, 23].
Известно, что во время выполнения мышечной работы до утомления отмечается накопление ионов H + и продуктов метаболитов [38, 39], сдвиг ионной концентрации Na + и K + [23]. Такие изменения могут нарушать связь возбуждение—сокращение, включающее свойства мышечной мембраны и генерацию ПД мышцы, ведущих в ЭМГ проявлений утомления мышцы, но отличающееся от механических проявлений [13, 27, 32]. Возможно, это не единственные причины и утомление может иметь место в каждом из перечисленных моментов. Все это указывает не только на связь между неупотреблением и изменением во внутриклеточных механизмах, контролирующих кинетику мышечного сокращения, но и предполагает, что изменение ионной проницаемости мышечной мембраны может существенно влиять на сократительную кинетику мышцы.
Цель настоящего исследования — определить паттерн утомления при ритмическом тетаническом сокращении ТМГ у человека после 7-суток гравитационной разгрузки. Механические и электрические параметры мышцы регистрировали во время электрической стимуляции ее моторного нерва, что позволяло отделить периферический аппарат от центрального управления. Сопоставление изменений в сократительных и электрических ответах этой группы мышц привлекает должное внимание со стороны многих исследователей и, в частности занимающихся проблемой космической физиологии.
МЕТОДИКА
Испытуемые и иммерсионная процедура
Функциональные свойства периферического нервно-мышечного аппарата (НМА) оценивали дважды — до и после механической разгрузки, и протокол исследований был идентичным.
Экспериментальная процедура
Работоспособность периферического НМА, на примере ТМГ, у человека в условиях in situ оценивали по крутизне снижения тетанического изометрического сокращения, продуцируемого ТМГ при ее ритмическом тетаническом сокращении. Сокращение мышцы вызывали супрамаксимальными электрическими импульсами, используя монополярный электрод, помещенный накожно в подколенной ямке на область n. tibialis. Утомление исследовали при выполнении 2 мин стандартной серии, состоящей из изометрических ритмических 1 с сокращений, разделенных паузой отдыха той же длительности [5]. Частота стимуляции 50 имп/с, физиологическая частота, с одной стороны, близкая к частоте активации мышечных клеток во время начального развития сильного произвольного сокращения [15], а с другой — позволяет развивать максимальную силу сокращения мышцы 2.
Стимуляция
N. tibialis электрически стимулировали прямоугольными супрамаксимальными (+40 %) импульсами длительностью каждого 1 мс, используя монополярный электрод (диаметром 1 см). Активный электрод (катод) устанавливали в подколенной ямке (область наименьшего сопротивления) на моторный нерв, пассивный (размером 6 х 4 см) — на нижней трети передней поверхности бедра. Электрические импульсы подавали от нейромышечного стимулятора (тип ЭСУ-1, СССР) через изолирующую приставку. Использовали одиночные и тетанические ритмические импульсы.
Регистрация механических и электрических ответов
Регистрацию электрической активности мышцы, электрический сигнал или поверхностный ПД (ППД), осуществляли поверхностными биполярными AG-AgCl чашечными (8 мм) электродами, расположенными на уровне ниже 6 см головок m. gastrocnemius по средней линии брюшка m. soleus, с межэлектродным расстоянием 25 мм. Положение отводящих электродов подбирали таким образом, чтобы форма электрических ответов мышцы — М-ответов, была наиболее простой. Большой электрод заземления (размером 7.5 х 6.5 см) устанавливали в проксимальной части голени между отводящими и регистрирующими электродами. ППД усиливали усилителем (тип УБП-1-02, СССР), а затем синхронно осуществляли запись на светолучевом осциллографе и, для визуального контроля, на экране запоминающего осциллоскопа (тип С8-9А, СССР).
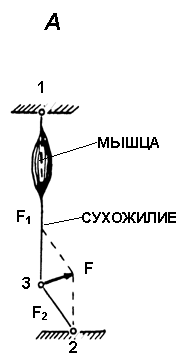
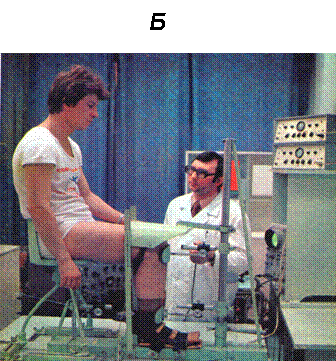
Рис. 1.
А. Схема регистрации силы сокращения скелетной мышцы у человека с помощью тендометрического датчика в условиях in situ.
Б. Тендометрическая установка и позиция человека при регистрации механических и электрических ответов отдельной скелетной мышцы голеностопного сустава.
Регистрацию механических и электрических ответов мышцы во время ритмической тетанической стимуляции осуществляли на протяжении 1 с после начала тетанизации нерва, а затем короткими (около 0.2 с) отрезками в конце каждого сокращения.
Электрические и механические ответы ТМГ регистрировали на специальной ленте светолучевого осциллографа и дополнительно на экране запоминающего осциллографа.
Измерение
По тендограмме изометрического одиночного сокращения мышцы, в ответ на одиночный электрический импульс n. tibialis, измеряли силу (пик амплитуды) одиночного сокращения (Рос), время одиночного сокращения (ВОС) и время полурасслабления (1/2 ПР) мышцы.
Рассчитывали также максимальную скорость (dPoc/dt) развития изометрического одиночного сокращения.
Статистика
При обработке полученных результатов исследования рассчитывали среднюю и стандартную ошибку средней (М ± m). Различие между фоновыми (контрольными) показателями и показателями, зарегистрированными после АНОГ, оценивали с помощью параметрического t-критерия Стьюдента и непараметрического критерия Вилкоксона. Уровень достоверности р 0.05).
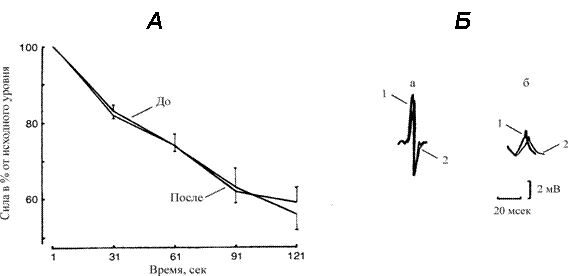
Рис. 2.
А. Изменение силы сокращения трехглавой мышцы голени во время 60 ритмических 1‑с электрически вызванных (50 имп/с) сокращений с 1‑с интервалами отдыха между сокращениями в до и после 7 суточной механической разгрузки. Величины выражены как % от исходной тетанической силы сокращения.
Б. Поверхностный потенциал действия, зарегистрированный в начале (а) и на
60 ритмическом сокращении (б) в контрольных условиях (1) и после разгрузки (2).
Вместе с тем, анализ кривых изменения сократительной активности мышцы обнаружил в основном бифазный характер ответа. При этом каждая последующая фаза характеризуется своей динамикой изменения силы сокращения мышцы. Более быстрое снижение силы сокращения от исходной величины отмечается на 30-31 с (17-18 %) с последующим относительным замедлением к концу 60-61 с (8-9 %) и вновь некоторым возрастанием скорости снижения силы сокращения отмечается к 90-91 с (11-12 %) и с замедлением на последнем участке кривой (3-7 %). Это наблюдение кажется парадоксальным, поскольку хорошо известно, что во время сильных изометрических сокращений механическое напряжение мышцы препятствует внутримышечной циркуляции кровотока и развивается ишемический процесс [20]. Результат предполагает, что эти две фазы утомления обусловлены разными процессами — по возможности чувствительностью к внутримышечному циркулирующему кровотоку.
Интересно, что после механической разгрузки не обнаруживаются различий в кривой снижения мышечного напряжения при выполнении теста на утомление. Наше наблюдение, что развиваемая во время сокращения мышцы ишемия видоизменяет первоначальную фазу соотношения между временем сокращения и уровнем снижения силы, и значительно ослабевает во второй части кривой, допускает, что разные механизмы вовлекаются в двух фазах.
Утомление и электрические процессы
Уменьшение продуцируемой механической силы сокращения может быть результатом снижения электрической активности мышцы. В этой связи мы проанализировали ЭМГ активность мышцы (см. раздел «МЕТОДИКА») и усредненные данные представлены в табл. 1 и графически на рис. 2, Б. Наши результаты показывают, что изменение электрических процессов, наблюдаемых во время выполнения ритмических тестовых сокращений на утомление, различается в контроле и после ее механической разгрузки. После 7-суточной разгрузки мышечного аппарата отмечается уменьшение амплитуды негативной фазы ПД и увеличение длительности М-ответа. По существу эти изменения начинаются с первого М-ответа и возрастают в конце 1, 5 и особенно на 121 с тестового сокращения (табл. 1; рис. 2-Б).
Таблица 1. Электрические свойства мышцы до и после 7-суточной механической разгрузки
Поверхностный потенциал действия
Время от момента начала электрической стимуляции, с
Как показывают наши данные, снижение силы сокращения мышцы в тесте на утомление не отличалось в контроле и после разгрузки, указывая, с одной стороны, что развиваемое утомление нельзя объяснить изменениями в самом сократительном звене НМА в результате гравитационной разгрузки и, во-вторых, что одним из компонентов (если не единственной причиной), развивающегося периферического утомления может быть нарушение в «электрогенном» звене НМА. Сопоставление изменений сократительного ответа с соответствующим изменением электрического ответа (Э/С-отношение) позволяет определить удельную роль «электрогенного» и сократительного звеньев в развитии периферического утомления. Анализ Э/С-отношения до и после разгрузки обнаружил, что возникает неодинаковая динамика изменений в состоянии сократительного и электрического звеньев периферического НМА (см. рис. 4). Эти результаты не противоречат вышеописанной дискуссии, поскольку трактовка Э/С‑отношения предполагает линейную связь между величиной электрического и механического ответов мышцы. Однако, последняя имеет нелинейную связь [27], и особенно при утомлении может нарушаться. При вызванном сокращении, триггером которого является электрическая стимуляция моторного нерва, уменьшение силы мышечного волокна может компенсироваться модуляцией частоты разряда двигательных единиц, не оказывая существенного влияния на силу тетанического сокращения мышечного волокна [40]. Кроме того, нормальный ПД мышечного волокна в несколько раз превышает пороговую величину, необходимую для активации сократительного аппарата [34], в результате даже уменьшенные ПД способны запускать нормальное сокращение мышечных волокон.
ЗАКЛЮЧЕНИЕ
Утомление, развивающееся при выполнении упражнений короткой длительности (как в настоящем исследовании) и, проявляющееся в снижение тетанической силы сокращения мышцы, является следствием изменений периферического НМА. Исследование электрически вызванного сокращения вносит существенный вклад в понимание основных механизмов периферического утомления НМА, поскольку позволяет исключить влияние модуляции частоты импульсации моторных, двигательных, единиц на регистрируемые параметры механических ответов мышцы.
Результаты настоящего исследования показывают, что механическое ослабление изометрического напряжения во время ритмических мышечных сокращений определяется периферическими факторами и механизмы ассоциируются с мембраной и внутриклеточными процессами. Сравнение снижения электрических ответов и механического ответа во время ритмических электрически вызванных сокращений в обоих тестовых сокращениях указывает, что замедление генерации ПД вдоль нервной и мышечной мембран, также как и возможно увеличение синаптической задержки, не объясняет ослабления развиваемого напряжения. Сравнение электрических и механических ответов, зарегистрированных в контрольных условиях и после гравитационной разгрузки, поддерживает предположение, что электрические изменения не тесно контролируют механическое ослабление. Таким образом, нервно-мышечные изменения, наблюдаемые во время выполнения короткой длительности упражнений, внутриклеточные процессы могут играть доминирующую роль в отношении наблюдаемого снижения силы сокращения во время выполнения дозированного теста на утомление после функциональной разгрузки.
Благодарность
Автор выражает также особую благодарность и признательность всем испытуемым, принявшим участие в эксперименте, без которых невозможно было бы получить фактический материал.
Нормальная физиология (7 стр.)

Если каждый последующий стимул поступает к мышце в тот период, когда она находится в фазе укорочения, то возникает гладкий тетанус, а если в фазу расслабления – зубчатый тетанус.
Амплитуда тетанического сокращения превышает амплитуду одиночного мышечного сокращения. Н. Е. Введенский объяснил это явление фазными изменениями возбудимости мышцы, введя понятие об оптимуме и пессимуме частоты раздражения. Оптимум – такая частота раздражения, при которой каждое последующее раздражение наносится в фазу повышенной возбудимости. Тетанус при этом будет максимальным по амплитуде. Пессимум – такая частота раздражения, при которой каждое последующее раздражение наносится в фазу пониженной возбудимости. Амплитуда тетануса при этом будет минимальной.
Различают несколько видов мышечных сокращений: изотонический, изометрический и смешанный. При изотоническом сокращении мышцы происходит изменение ее длины, а напряжение остается постоянным. Такое сокращение происходит в том случае, если отсутствует сопротивление изменению ее длины. К изотоническому типу сокращений относятся сокращения мышц языка. При изометрическом сокращении длина мышечных волокон остается постоянной, а их напряжение возрастает. Такое сокращение мышцы возникает при попытке поднять чрезмерно большой груз. В естественных условиях сокращения мышц никогда не бывают чисто изотоническими или изометрическими, они имеют смешанный характер, т. е. происходит изменение и длины, и напряжения мышцы.
Механизм мышечного сокращения
Мышцы состоят из мышечных волокон, а те – из множества тонких нитей – миофибрилл, расположенных продольно. Каждая миофибрилла состоит из нитей сократительных белков актина и миозина. Перегородки, называемые Z-пластинами, разделяют миофибриллы на участки – саркомеры. В саркомере чередуются поперечные светлые и темные полосы. Поперечная исчерченность миофибрилл обусловлена определенным расположением нитей актина и миозина. В центральной части каждого саркомера расположены толстые нити миозина. На обоих концах саркомера находятся тонкие нити актина, прикрепленные к Z-пластинам. Нити миозина выглядят в световом микроскопе как светлая полоска (Н-зона) в темном диске, который содержит нити миозина и актина и называется анизотропным, или А-диском. По обе стороны от А-диска находятся участки, которые содержат только тонкие нити актина и кажутся светлыми, они называются изотропными, или I-дисками. По их середине проходит темная линия – Z-мембрана. Благодаря такому периодическому чередованию светлых и темных дисков сердечная и скелетная мышцы выглядят поперечно-полосатыми (рис.7).
В состоянии покоя концы толстых и тонких нитей лишь незначительно перекрываются на уровне А-диска. При сокращении тонкие актиновые нити скользят вдоль толстых миозиновых нитей, двигаясь между ними к середине саркомера. Сами актиновые и миозиновые нити своей длины не изменяют. Миозиновые нити имеют поперечные мостики (выступы) с головками, которые отходят от нити биполярно. Актиновая нить состоит из двух закрученных одна вокруг другой цепочек молекул актина. На нитях актина расположены молекулы тропонина, а в желобках между двумя актина лежат нити тропомиозина. Молекулы тропомиозина в покое располагаются так, что предотвращают прикрепление поперечных мостиков миозина к актиновым нитям.

Во многих местах участки поверхностной мембраны мышечной клетки углубляются в виде трубочек внутрь волокна, перпендикулярно его продольной оси, образуя систему поперечных трубочек (Т-систему). Параллельно миофибриллам и перпендикулярно поперечным трубочкам расположена система продольных трубочек (альфа-система). Пузырьки на концах этих трубочек, в которых сосредоточено основное количество внутриклеточного кальция, подходят очень близко к поперечным трубочкам, образуя совместно с ними так называемые триады. В состоянии покоя миозиновый мостик заряжен энергией (миозин фосфорилирован), но он не может соединиться с нитью актина, так как между ними находится система из нитей тропомиозина и молекул тропонина. При возбуждении ПД распространяется по мембранам Тсистемы внутрь клетки и вызывает высвобождение ионов кальция из альфа-системы. С появлением ионов кальция в присутствии АТФ происходит изменение пространственного положения тропонина – нить тропомиозина сдвигается и открываются участки актина, присоединяющие миозиновые головки. Соединение головки фосфорилированного миозина с актином приводит к изменению положения мостика (его «сгибанию»), в результате нити актина перемещаются на 1 мм к середине саркомера. Затем происходит отсоединение мостика от актина. Ритмические прикрепления и отсоединения головок миозина тянут актиновую нить к середине саркомера.
При отсутствии повторного возбуждения ионы кальция закачиваются кальциевым насосом из межфибриллярного пространства в систему саркоплазматического ретикулума. Это приводит к снижению концентрации ионов кальция и отсоединению его от тропонина. Вследствие чего тропомиозин возвращается на прежнее место и снова блокирует активные центры актина. Затем происходит фосфорилирование миозина за счет АТФ, что также способствует временному разобщению нитей. Расслабление мышцы после ее сокращения происходит пассивно – актиновые и миозиновые нити легко скользят в обратном направлении под влиянием сил упругости мышечных волокон, а также сокращения мышц-антагонистов.
Гладкие мышцы
Эти мышцы образуют мышечные слои стенок желудка, кишечника, мочеточников, бронхов, кровеносных сосудов и других внутренних органов. Они построены из веретенообразных одноядерных мышечных клеток. Гладкие мышцы разделяются на две основные группы: мультиунитарные и унитарные. Мультиунитарные мышцы функционируют независимо друг от друга, и каждое волокно может иннервироваться отдельным нервным окончанием. Такие волокна обнаружены в ресничной мышце глаза, мигательной перепонке и мышечных слоях некоторых крупных сосудов, к ним относятся мышцы, поднимающие волосы. У унитарных мышц волокна настолько тесно переплетены, что их мембраны могуг сливаться, образуя электрические контакты (нексусы). При раздражении одного волокна за счет этих контактов ПД быстро распространяются на соседние волокна. Поэтому, несмотря на то, что двигательные нервные окончания расположены на небольшом числе мышечных волокон, в реакцию вовлекается вся мышца. Такие мышцы имеются в большинстве органов: пищеварительном тракте, матке, в мочеточниках.
Особенностью гладких мышц является их способность осуществлять медленные и длительные тонические сокращения. Медленные, ритмические сокращения гладких мышц желудка, кишечника, мочеточников и других органов обеспечивают перемещение содержимого этих органов. Длительные тонические сокращения гладких мышц обеспечивают функционирование сфинктеров полых органов, которые препятствуют выходу их содержимого.
Гладкие мышцы стенок кровеносных сосудов, особенно артерий и артериол, также находятся в состоянии постоянного тонического сокращения. Изменение тонуса мышц стенок артериальных сосудов влияет на величину их просвета и, следовательно, на уровень кровяного давления и кровоснабжения органов. Важным свойством гладких мышц является их пластичность, т. е. способность сохранять приданную им при растяжении длину. Скелетная мышца в норме почти не обладает пластичностью. При удалении растягивающего груза скелетная мышца быстро укорачивается, а гладкая остается растянутой. Высокая пластичность гладких мышц имеет большое значение для нормального функционирования полых органов. Например, пластичность мышц мочевого пузыря по мере его наполнения предотвращает избыточное повышение давления.
Сильное и резкое растяжение гладких мышц вызывает их сокращение, что обусловлено нарастающей при растяжении деполяризацией клеток, которая обеспечивает автоматию гладкой мышцы. Такое сокращение играет важную роль в авторегуляции тонуса кровеносных сосудов, а также способствует непроизвольному опорожнению переполненного мочевого пузыря в тех случаях, когда нервная регуляция отсутствует в результате повреждения спинного мозга.
В гладких мышцах тетаническое сокращение возникает при низкой частоте стимуляции. В отличие от скелетных, гладкие мышцы способны развивать спонтанные тетанообразные сокращения в условиях денервации и даже после блокады интрамуральных ганглиев. Такие сокращения возникают вследствие активности клеток, обладающих автоматией (пейсмекерных клеток), которые отличаются по электрофизиологическим свойствам от других мышечных клеток. В них появляются пейсмекерные потенциалы, деполяризующие мембрану до критического уровня, что вызывает возникновение потенциала действия.
Особенностью гладких мышц является их высокая чувствительность к медиаторам, которые оказывают на спонтанную активность пейсмекеров модулирующие влияния. При нанесении ацетилхолина на препарат мышцы толстой кишки частота ПД возрастает. Вызываемые ими сокращения сливаются, образуется почти гладкий тетанус. Чем выше частота ПД, тем сильнее сокращение. Норадреналин, напротив, гиперполяризует мембрану, снижая частоту ПД и величину тетануса.
Возбуждение гладкомышечных клеток вызывает повышение концентрации кальция в саркоплазме, что активирует сократительные структуры. Так же как сердечная и скелетная мышцы, гладкая мышца расслабляется при снижении концентрации ионов кальция. Расслабление гладких мышц происходит медленнее, так как удаление ионов кальция замедлено.
Тетанус
Тетаническое сокращение
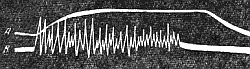 | Рис. 142. Одновременная запись тетанического сокращения (А) и токов действия (В) мышцы (по А. Г. Гинецинскому). |
Тетанические сокращения мышцы представля ют собой результат суммации одиночных сокращений. Для исследования суммации сокращений наносят на мышцу два одиночных раздражения. Промежуток времени между раздражениями должен быть такой, чтобы второе раздражение подействовало на мышцу раньше, чем она успеет расслабиться после первого сокращения. При этом возможны два случая.
Если второе раздражение поступает, когда мышца уже начала расслабляться после первого сокращения, то на миографической кривой вершина второго сокращения будет отделена от вершины первого небольшим западением кривой ( рис. 143, Б ). Если же второе раздражение действует, когда первое сокращение ещё не дошло до своей вершины, то второе сокращение полностью сливается с первым, образуя вместе с ним единую суммированную вершину ( рис. 143, А ).
 |
 |
 |