Пролиферация клеток что это
В развитии низших и высших позвоночных отчетливо прослеживается единая общебиологическая закономерность, выражающаяся в появлении зародышевых листков и обособлении основных зачатков органов и тканей. Процесс образования тканей из материала эмбриональных зачатков составляет суть учения о гистогенезах.
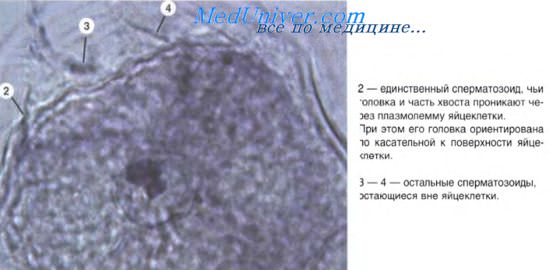
Эмбриональный гистогенез, по определению А.А. Клишова (1984), — это комплекс координированных во времени и пространстве процессов пролиферации, клеточного роста, миграции, межклеточных взаимодействий, дифференциации, детерминации, программированной гибели клеток и некоторых других. Все названные процессы в той или иной мере протекают в зародыше, начиная с самых ранних стадий его развития.
Пролиферация. Основной способ деления тканевых клеток — это митоз. По мере увеличения числа клеток возникают клеточные группы, или популяции, объединенные общностью локализации в составе зародышевых листков (эмбриональных зачатков) и обладающие сходными гистогенетическими потенциями. Клеточный цикл регулируется многочисленными вне- и внутриклеточными механизмами. К внеклеточным относятся влияния на клетку цитокинов, факторов роста, гормональных и нейрогенных стимулов. Роль внутриклеточных регуляторов играют специфические белки цитоплазмы. В течение каждого клеточного цикла существуют несколько критических точек, соответствующих переходу клетки из одного периода цикла в другой. При нарушении внутренней системы контроля клетка под влиянием собственных факторов регуляции элиминируется апоптозом, либо на некоторое время задерживается в одном из периодов цикла.
Метод радиографического анализа клеточных циклов в различных тканях выявил особенности соотношения клеточной репродукции и дифференцировки. Например, если в тканях (кроветворные ткани, эпидермис) имеется постоянный фонд пролиферирующих клеток, за счет которых обеспечивается непрерывное возникновение новых клеток взамен погибающих, то эти ткани относятся к обновляющимся. Другие ткани, например, некоторые соединительные, характеризуются тем, что в них увеличение количества клеток происходит параллельно с их дифференцировкой, клетки в этих тканях характеризуются низкой митотической активностью. Это растущие ткани. Наконец, нервная ткань характеризуется тем, что все основные процессы репродукции заканчиваются в период эмбрионального гистогенеза (когда формируется основной запас стволовых клеток, достаточный для последующего развития ткани). Поэтому она отнесена к стабильным (стационарным) тканям. Продолжительность жизни клеток в обновляющихся, растущих и стабильных тканях разная.
Наряду с обновлением клеточной популяции, в самих клетках постоянно наблюдается обновление внутриклеточных структур (внутриклеточная физиологическая регенерация).
Клеточный рост, миграция и межклеточные взаимодействия. Рост клеток проявляется в изменении их размеров и формы. При усилении функциональной активности и внутриклеточных биосинтезов наблюдается увеличение объема клетки. Если объем клетки превышает некую норму, то говорят о ее гипертрофии, и наоборот, при снижении функциональной активности происходит уменьшение объема клетки, а при переходе некоторых нормативных параметров возникает атрофия клетки. Рост клетки не беспределен и определяется оптимальным ядерно-цитоплазменным отношением.
Важное значение для гистогенеза имеют процессы перемещения клеток. Миграция клеток наиболее характерна для периода гаструляции. Однако и в период гисто- и органогенеза происходят перемещения клеточных масс (например, смещения миобластов из миотомов в места закладки скелетных мышц; движение клеток из нервного гребня с образованием спинномозговых ганглиев и нервных сплетений, миграция гоноцитов и т. д.). Миграция осуществляется с помощью нескольких механизмов. Так, различают хемотаксис — движение клеток в направлении градиента концентрации какого-либо химического агента (перемещения спермиев к яйцеклетке, предшественников Т-лимфоцитов из костного мозга в закладку тимуса).
Гаптотаксис — механизм перемещения клеток по градиенту концентрации адгезионной молекулы (движение клеток протока пронефроса у амфибий по градиенту щелочной фосфатазы на поверхности мезодермы). Контактное ориентирование — когда в какой-либо преграде остается один канал для перемещения (описан у рыб при образовании плавников).
Контактное ингибирование — этот способ перемещения наблюдается у клеток нервного гребня. Суть способа заключается в том, что при образовании ламеллоподии одной клеткой и контакта ее с другой клеткой, ламеллоподия прекращает рост и постепенно исчезает, но в другой части мигрирующей клетки при этом формируется новая ламеллоподия.
В процессе миграции клеток важную роль играют межклеточные взаимодействия. Существует несколько механизмов такого взаимодействия (контактного и дистантного). Выделяется большая группа молекул клеточной адгезии (МКА). Так, кадгерины — это Са2+-зависимые МКА, отвечают за межклеточные контакты при образовании тканей, за формообразование и др. В молекуле кадгерина различают внеклеточный, трансмембранный и внутриклеточный домены. Например, внеклеточный домен ответственен за адгезию клеток с одинаковыми кадгеринами, а внутриклеточный — за форму клетки. Другой класс МКА — это иммуноглобулиновое суперсемейство Са2+-независимых МКА, обеспечивающих, например, адгезию аксонов к сарколемме мышечных волокон, или миграцию нейробластов вдоль радиальных глиоцитов в закладке коры большого мозга и др. Следующий класс МКА — это мембранные ферменты — гликозилтранферазы. Последние по типу «ключ-замок» соединяются с углеводными субстратами — гликозаминогликанами надмембранного комплекса клетки, осуществляя таким образом прочное сцепление клеток.
Кроме механизмов межклеточного взаимодействия, существуют механизмы взаимодействия клеток с субстратом. Они включают формирование рецепторов клетки к молекулам внеклеточного матрикса. К последним относят производные клеток, среди которых наиболее изученными адгезионными молекулами являются коллаген, фибронектин, ламинин, тенасцин и некоторые другие. Коллагены, среди которых различают несколько десятков типов, входят в состав межклеточного вещества рыхлой волокнистой соединительной ткани, базальной мембраны и пр. Фибронектин, секретируемый клетками, является связывающей молекулой между мигрирующей клеткой и межклеточным матриксом. Ламинин — компонент базальной мембраны, также связывает мигрирующие клетки с межклеточным матриксом (справедливо по отношению к эпителиоцитам и нейробластам).
Для осуществления связи мигрирующих клеток с межклеточным матриксом клетки формируют специфические рецепторы. К ним относятся, например, синдекан, который обеспечивает контакт эпителиоцита с базальной мембраной за счет сцепления с молекулами фибронектина и коллагена. Интегрины клеточных поверхностей связывают с внеклеточной стороны молекулы внеклеточного матрикса, а внутри клетки — белки цитоскелета (например, актиновые микрофиламенты). Так возникает связь внутри- и внеклеточных структур, что позволяет клетке использовать для перемещения собственный сократительный аппарат. Наконец, существует большая группа молекул, формирующих клеточные контакты, осуществляющие коммуникацию между клетками (щелевые контакты), механическую связь (десмосомы, плотные контакты).
Дистантные межклеточные взаимодействия осуществляются путем секреции гормонов и факторов роста (ФР). Последние — это вещества, оказывающие стимулирующее влияние на пролиферацию и дифференцировку клеток и тканей. К ним относятся, например, ФР, полученный из тромбоцитов и влияющий на переход клеток в фазу размножения (гладких миоцитов, фибробластов, глиоцитов); эпидермальный ФР — стимулирует пролиферацию эпителиоцитов, производных эктодермы; ФР фибробластов — стимулирует пролиферацию фибробластов. Особо выделяется большая группа пептидов (соматотропины, соматомедины, инсулин, лактоген), влияющих на развитие клеток плода.
Факторы, тормозящие пролиферацию и дифференцировку клеток, также принимают кооперативное участие в процессах развития клеток и тканей. К ним относятся, например, бета-интерферон и трансформирующий ФР. Последний, однако, в отношении разных клеточных типов действует по-разному: блокирует размножение многих типов эпителиоцитов, но стимулирует размножение соединительнотканных клеток.
— Вернуться в оглавление раздела «гистология»
Пролиферация клеток что это
Пролиферативная активность клеток отражает интенсивность роста опухолей. В статье приведены данные изучения уровня пролиферации клеток базалиомы — наиболее частой злокачественной опухоли кожи. Исследование проводили иммуногистохимическим методом с маркером пролиферации Ki67. По характеру пролиферации базалиома представляла гетерогенную группу. Солидный, аденоидный, поверхностный мультицентрический тип базалиомы отличались низкой пролиферативной активностью с преимущественным распределением клеток, экспрессирующих Ki67, по периферии комплексов. Высокую пролиферативную активность выявляли в инфильтративном, склерозирующем и микронодулярном типе при равномерном распределении пролиферирующих клеток.
Пролиферативная активность клеток отражает интенсивность роста опухолей. В статье приведены данные изучения уровня пролиферации клеток базалиомы — наиболее частой злокачественной опухоли кожи. Исследование проводили иммуногистохимическим методом с маркером пролиферации Ki67. По характеру пролиферации базалиома представляла гетерогенную группу. Солидный, аденоидный, поверхностный мультицентрический тип базалиомы отличались низкой пролиферативной активностью с преимущественным распределением клеток, экспрессирующих Ki67, по периферии комплексов. Высокую пролиферативную активность выявляли в инфильтративном, склерозирующем и микронодулярном типе при равномерном распределении пролиферирующих клеток.
В структуре злокачественных эпителиальных опухолей кожи первое место занимает базалиома, на долю которой приходится от 75 до 94% [1, 2]. В подавляющем большинстве случаев регистрируются солитарные опухоли, локализующиеся на открытых участках кожи, подверженных инсоляции. Опухоль отличается медленным течением и относительно благоприятным прогнозом. Однако в ряде случав она может значительно увеличиваться в размере, характеризуясь выраженным экзофитным ростом, либо приобретать инвазивные черты, проникая в окружающие и подлежащие ткани, приводя к их разрушению. Подобный рост опухоли обычно ассоциируется с определенными морфологическими типами, которые рассматривают как агрессивные [3].
Показателем интенсивности роста ткани является пролиферативная активность клеток. Она относительно спокойна в доброкачественных процессах и резко возрастает в злокачественных опухолях. Аналогичные тенденции присущи и эпителиальным опухолям кожи. Так, пролиферативная активность клеток базальноклеточного рака кожи выше, чем в очагах лейкоплакии [4], а при раке in situ индекс пролиферации превосходит таковой при предраковом актиническом кератозе [4,5]. Плоскоклеточный рак кожи отличается высоким уровнем пролиферации, который следует за снижением степени его дифференцировки [6, 7, 8].
Предыдущие исследования показали, что по характеру пролиферативной активности клеток базалиома представляет гетерогенную группу. Наиболее интенсивный рост имела язвенная форма, тогда как нодулярная и поверхностная характеризовались умеренными и низкими показателями [9]. Высокий уровень пролиферации отмечали в микронодулярном, метатипическом типе базалиомы и типе морфеа [10, 12]. Также отмечали повышение индекса Ki67 в инфильтративном и метатипическом типе по сравнению с нодулярным и поверхностным [11, 13]. В то же время единого мнения о связи уровня пролиферации с гистологическим типом базалиомы до сих пор нет, так как другие исследования свидетельствовали, что индекс Ki67 в нодулярном и поверхностном мультицентрическом типе базальноклеточного рака значительно превосходил аналогичный показатель в склерозированном и типе морфеа [14, 15]. Учитывая противоречивые данные, представляется интересным и актуальным продолжить изучение пролиферативной активности клеток в различных морфологических типах базалиомы, что позволит более детально понимать маркеры и предикторы агрессивного течения опухоли.
Материалы и методы
В работе исследования использовали 36 биоптатов кожи больных базальноклеточным раком кожи. Биопсию кожи размером 0,5 х 0,5 см проводили под местной анестезией 2% раствором новокаина, биоптаты фиксировали в 10% растворе формалина, забуфференом по Лилли при рН-7,4. Материал заливали в парафин по обычной методике. Серийные срезы толщиной 3–5 мкм депарафинировали по стандартной схеме, затем окрашивали гематоксилином и эозином. Иммуногистохимическое исследование проводили в соответствии со стандартным протоколом. Парафиновые срезы наносили на стекла с адгезивным покрытием, депарафинировали по стандартному протоколу. Срезы, погруженные в цитратный буфер, инкубировали в микроволновой печи при мощности 690 Вт два раза по пять минут. Двадцать минут при комнатной температуре, срезы охлаждали, подсушивали и наносили первичные антитела. Для определения уровня пролиферации использовали моноклональные антитела к протеину Ki67 (DAKO), разведение 1:200. Использовались соответствующие позитивные и негативные контроли – иммунные и неиммунные сыворотки. Срезы промывали в буфере, подсушивали, затем на них наносили EnVision (anti-mouse и anti-rabbit, фирмы DAKO, Дания) на 30–40 минут. Для визуализации реакции наносили DAB+ (3, 3 ґ– диаминобензидин), что позволяло получать специфическую коричневую окраску. Затем срезы докрашивали гематоксилином Майера. Определение уровня пролиферации оценивалась с помощью индекса, который вычислялся как среднее от числа меченных ядер на 100 учтенных ядер (при учете 500–1000 опухолевых клеток). Подсчет меченных ядер проводился в репрезентативных полях зрения с относительно равномерным распределением опухолевых клеток, сверху вниз и слева направо. Клетки на периферии срезов, где чаще всего наблюдается фоновое окрашивание, не учитывались.
Среди 36 базалиом пять имели поверхностный мультицентрический тип строения, четыре – поверхностный мультицентрический в сочетании с солидным, семь – солидный, два – аденоидный, два – солидно-аденоидный, пять – инфильтративный, три – микронодулярный. В восьми случаях отмечали различные варианты сочетания солидного, инфильтративного, микронодулярного, склерозирующего строения.
Индекс пролиферации Ki67 в поверхностном мультицентрическом типе и при сочетании его с солидным компонентом варьировал от 14,9 до 31,1%. Средний индекс составлял 22,2%. Пролиферирующие клетки в шести случаях располагались преимущественно по периферии клеточных комплексов, в трех – равномерно по клеточным комплексам. При солидных опухолях средний индекс составил 26,5%, он варьировал от 14,5 до 38,2% (рис.1). Периферическое распределение наблюдали в четырех случаях, равномерное – в трех. При аденоидном строении пролиферирующие клетки распределялись равномерно, индекс пролиферации составлял 22,6 и 31,3%. В случаях, когда имелось сочетание солидного и аденоидного компонентов, индекс пролиферации составлял 36,8 и 19,2%, В первом случае клетки распределялись равномерно, во втором – равномерно в аденоидном компоненте и по периферии – в солидном. В опухолях инфильтративного, микронодулярного строения и при их сочетании клетки, экспрессирующие маркер Ki67, диффузно регистрировались по всем клеточным комплексам. В инфильтративном типе минимальное и максимальное значение индекса было 27,6 и 47,8% (среднее значение – 35,4%) (рис.2). Микронодулярные базалиомы имели средний индекс Ki67 26,9%. В случаях инфильтративно-склерозирующего строения индекс имел значение 40,2 и 31,6%, пролиферирующие клетки равномерно распределялись в комплексах (рис.3). При сочетании инфильтративного, солидного и микронодулярного строения преимущественно отмечали равномерное распределение пролиферирующих клеток, и средний индекс составлял 36,5% (значения варьировали от 22,1 до 50,4%). Солидное строение в сочетании с инфильтративным склерозирующим имели опухли с индексом Ki67 26,7 и 35,3%, в солидном компоненте пролиферирующие клетки располагались преимущественно по периферии комплексов, инфильтративном склерозирующем – равномерно.
При анализе пролиферативной активности клеток мы сгруппировали опухоли в зависимости от характера течения и роста. В группу неинвазивного роста мы включили опухоли, имеющие поверхностный мультицентрический, солидный, аденоидный, сочетание поверхностного мультицентрического и солидного компонентов. В группу с инвазивным ростом нами были включены опухоли инфильтративные, микронодулярные, инфильтративно склерозирующие, а также опухоли сложного строения (инфильтративный + микронодулярный + солидный и солидый + инфильтративный склерозирующий).
Средний индекс пролиферации неагрессивных типов составил 24,77 ± 7, 47 с доверительным интервалом 95% 21,27–28,26 (табл. 1). Средний индекс Ki67 агрессивных типов составлял 33,53 ± 8,94 с доверительным интервалом 95% 28,77–38,29 (табл. 1). Различия в уровне пролиферации между агрессивными и неагрессивными типами носили статистически значимый характер (р = 0,003). Анализ характера распределения пролиферирующих клеток в опухолевых комплексах показал, что в неагрессивных типах периферическое распределение клеток наблюдалось в 50% случаев, равномерное – в 45%, в 5% случаев ряд комплексов в опухоли имели один тип распределения, другие комплексы – иной тип распределения (табл.2). В агрессивных типах 81,25% случаев имели равномерное распределение пролиферирующих клеток, сочетание двух типов распределения – 18,75% (табл. 2). Периферическое распределение пролиферирующих клеток в агрессивных типах не встречалось. При сравнении распределения клеток между агрессивными и неагрессивными типами опухоли было показано, что различия носят статистически достоверный характер (р = 0,003).
Таблица 1. Индекс пролиферации Ki67 в агрессивных и неагрессивных морфологических типах базалиомы
Научная электронная библиотека

Попков В. М., Чеснокова Н. П., Ледванов М. Ю.,
3.8. Механизмы развития пролиферации в очаге воспаления
Пролиферация является завершающей фазой развития воспаления, обеспечивающей репаративную регенерацию тканей на месте очага альтерации. Пролиферация развивается с самого начала воспаления наряду с явлениями альтерации и экссудации.
При репаративных процессах в очаге воспаления регенерация клеток и фиброплазия достигаются как активацией процессов пролиферации, так и ограничением апоптоза клеток. Размножение клеточных элементов начинается по периферии зоны воспаления, в то время как в центре очага могут еще прогрессировать явления альтерации и некроза. Полного развития пролиферация соединительнотканных и органоспецифических клеточных элементов достигает после «очистки» зоны повреждения от клеточного детрита и инфекционных возбудителей воспаления тканевыми макрофагами и нейтрофилами. В связи с этим следует отметить, что процессу пролиферации предшествует формирование нейтрофильного и моноцитарного барьеров, которые формируются по периферии зоны альтерации.
Восстановление и замещение поврежденных тканей начинается с выхода из сосудов молекул фибриногена и образования фибрина, который формирует своеобразную сетку, каркас для последующего клеточного размножения. Уже по этому каркасу распределяются в очаге репарации быстро образующиеся фибробласты. Деление, рост и перемещение фибробластов возможны только после их связывания с фибрином или коллагеновыми волокнами. Эта связь обеспечивается особым белком – фибронектином. Размножение фибробластов начинается по периферии зоны воспаления, обеспечивая формирование фибробластического барьера. Хемотаксис, активация и пролиферация фибробластов осуществляются под воздействием:
1. Факторов роста фибробластов.
2. Тромбоцитарного фактора роста.
3. Цитокинов – ФНО, ИЛ-1.
6. Трансформирующего фактора роста b.
Сначала фибробласты незрелые и не обладают достаточной синтетической активностью. Созреванию предшествует внутренняя структурно-функциональная перестройка фибробластов: гипертрофия ядра и ядрышка, гиперплазия ЭПС, повышение содержания ферментов, особенно щелочной фосфатазы, неспецифической эстеразы, b-глюкуронидазы. Только после перестройки фибробласты начинают синтезировать коллаген, эластин, коллагенассоциированные белки и протеогликаны. Коллагеногенез стимулируется следующими биологически активными веществами – ФНО, ИЛ-1, ИЛ-4, фактором роста фибробластов, тромбоцитарным фактором роста.
Интенсивно размножающиеся фибробласты продуцируют кислые мукополисахариды – основной компонент межклеточного вещества соединительной ткани (гиалуроновую кислоту, хондроитинсерную кислоту, глюкозамин, галактозамин). При этом зона воспаления не только инкапсулируется, но и начинаются постепенная миграция клеточных и бесклеточных компонентов соединительной ткани от периферии к центру, формирование соединительнотканного остова на месте первичной и вторичной альтерации.
Наряду с фибробластами размножаются и другие тканевые и гематогенные клетки. При разрушении базальных мембран сосудов в зоне альтерации происходит миграция клеток эндотелия по градиенту ангиогенных факторов. Просвет новообразующегося капилляра формируется путем слияния внеклеточных пространств соседних эндотелиоцитов. Вокруг новообразующихся капилляров концентрируются тучные клетки, макрофаги, нейтрофилы, которые освобождают биологически активные вещества, способствующие пролиферации капилляров.
Важнейшими факторами, стимулирующими ангиогенез, являются:
1. Факторы роста фибробластов (основной и кислый).
2. Сосудистый эндотелиальный фактор роста.
4. Эпидермальный фактор роста.
Фибробласты вместе с вновь образованными сосудами создают грануляционную ткань. Это, по существу, молодая соединительная ткань, богатая клетками и тонкостенными капиллярами, петли которых выступают над поверхностью ткани в виде гранул.
Основными функциями грануляционной ткани являются: защитная – предотвращение влияния факторов окружающей среды на очаг воспаления и репаративная – заполнение дефекта и восстановление анатомической и функциональной полноценности поврежденных тканей.
Формирование грануляционной ткани не строго обязательно. Это зависит от величины и глубины повреждения. Грануляционная ткань обычно не развивается при заживлении ушибленных кожных ранок или мелких повреждений слизистой оболочки [29]. Грануляционная ткань постепенно превращается в волокнистую ткань, называемую рубцом. В рубцовой ткани уменьшается количество сосудов, они запустевают, уменьшается количество макрофагов, тучных клеток, снижается активность фибробластов. Небольшая часть клеточных элементов, располагающаяся среди коллагеновых нитей, сохраняет активность. Предполагают, что сохранившие активность тканевые макрофаги принимают участие в рассасывании рубцовой ткани и обеспечивают формирование более мягких рубцов.
Параллельно с созреванием грануляций происходит эпителизация раны. Она начинается в первые часы после повреждения, и уже в течение первых суток образуются 2–4 слоя клеток базального эпителия. Скорость эпителизации обеспечивается следующими процессами: миграцией, делением и дифференцировкой клеток. Эпителизация небольших ран осуществляется, в основном, за счет миграции клеток из базального слоя. Раны более крупные эпителизируются за счет миграции и митотического деления клеток базального слоя, а также дифференцировки регенерирующего эпидермиса. Новый эпителий образует границу между поврежденным и подлежащим слоем, он препятствует обезвоживанию тканей раны, уменьшению в ней электролитов и белков, а также предупреждает инвазию микроорганизмов.
В процессе пролиферации участвуют и органспецифические клеточные элементы органов и тканей. С точки зрения возможностей пролиферации органспецифических клеточных элементов все органы и ткани могут быть расклассифицированы на три группы:
К первой группе могут быть отнесены органы и ткани, клеточные элементы которых обладают активной или практически неограниченной пролиферацией, достаточной для полного восполнения дефекта структуры в зоне воспаления (эпителий кожи, слизистых оболочек дыхательных путей, слизистой желудочно-кишечного тракта, мочеполовой системы; гемопоэтическая ткань и др.).
Ко второй группе относятся ткани с ограниченными регенерационными способностями (сухожилия, хрящи, связки, костная ткань, периферические нервные волокна).
К третьей группе относятся те органы и ткани, где органоспецифические клеточные элементы не способны к пролиферации (сердечная мышца, клетки ЦНС). Основными факторами, регулирующими процессы пролиферации и дифференцировки клеток в очаге воспаления, являются:
1. Факторы роста, продуцируемые макрофагами, лимфоцитами, тромбоцитами, фибробластами и другими клетками, стимулированными в зоне воспаления. К ним относятся:
? факторы роста эпидермиса (стимулятор пролиферации и созревания эпителия, стимулятор ангиогенеза);
? трансформирующий фактор роста-? (стимулятор ангиогенеза);
? трансформирующий фактор роста-? (хемоаттрактант фибробластов, стимулятор синтеза коллагена, фибронектина, ангиогенеза, ингибитор протеолиза);
? тромбоцитарный фактор роста (стимулятор миграции, пролиферации и синтеза белка в клетках-мишенях, обладает провоспалительным эффектом);
? фактор роста эндотелиоцитов;
? фактор роста фибробластов кислый и основной (стимуляторы пролиферации всех клеток сосудистой стенки);
фактор роста нервов (стимулятор пролиферации, роста, морфогенеза симпатических нейронов, эпителиальных клеток). Ростовые факторы, взаимодействуя с рецепторами на клетках- мишенях, могут непосредственно стимулировать синтез ДНК в клетках или подготавливать внутриклеточные рецепторы и ферменты к митотической деятельности.
2. Пептид гена, родственного кальцитонину, стимулирует пролиферацию эндотелиальных клеток, а субстанция Р индуцирует выработку ФНО в макрофагах.
3. Простагландины группы Е потенцируют регенерацию путем усиления кровоснабжения.
4. Кейлоны и антикейлоны, продуцируемые различными клетками, действуя по принципу обратной связи, могут активировать и угнетать митотические процессы в очаге воспаления [2].
5. Полиамины (путресцин, спермидин, спермин), обнаруживаемые во всех клетках млекопитающих, жизненно необходимы для роста и деления клеток.
Они обеспечивают стабилизацию плазматических мембран и суперспиральной структуры ДНК, защиту ДНК от действия нуклеаз, стимуляцию транскрипции, метилирование РНК и связывание ее с рибосомами, активацию ДНК-лигаз, эндонуклеаз, протеинкиназ и многие другие клеточные процессы. Усиленный синтез полиаминов, способствующих пролиферативным процессам, отмечается в очаге альтерации [5].
6. Циклические нуклеотиды: цАМФ ингибирует, а цГМФ активирует процессы пролиферации.