Вопрос 4. Полярность эпителиоцитов наиболее ярко проявляется в однослойных эпителиях




![]()
![]()
Вопрос 3
В многослойных эпителиях полярность проявляется в морфологической неоднородности клеток разных слоев.
Апикальная поверхность может образовывать ряд специальных структур:
– разновидностью микроворсинок считают стереоцилии и волоски рецепторных клеток внутреннего уха. Они гораздо крупнее, также поддерживаются изнутри актиновым цитоскелетом. При отклонении волосков мембрана слуховых клеток деполяризуется.
Базальная поверхность эпителиальных клеток чаще всего плоская. Однако у некоторых клеток образует складки, между которыми лежат митохондрии (базальная исчерченность). Такие эпителии активно перекачивают ионы из каких-либо жидкостей (например, процесс обратного всасывания мочи в почечных канальцах). Базальная сторона клеток всегда связана с базальной мембраной специальными контактами – полудесмосомами.
Базальная мембрана – это слой межклеточного вещества толщиной 20-100 нм и сложного белкового и полисахаридного состава. (коллаген IV типа, фибронектин, ламинин, гликозаминогликаны). Эти вещества определяют адгезию, упругость, проницаемость, коллоидное состояние, электрический заряд и другие свойства базальной мембраны. В составе базальной мембраны выделяют светлую пластинку, к которой непосредственно крепятся эпителиоциты и темную пластинку, в которую вплетены петли якорных коллагеновых фибрилл. Клетки прикреплены к базальной мембране специальными структурами – полудесмосомами. По общей организации они напоминают половинку десмосомы, но набор белков несколько иной. От прикрепительных пластинок в светлую пластинку базальной мембраны тянутся тонкие якорные филаменты. Со стороны соединительной ткани в петли якорных фибрилл закреплены коллагеновые фибриллы.
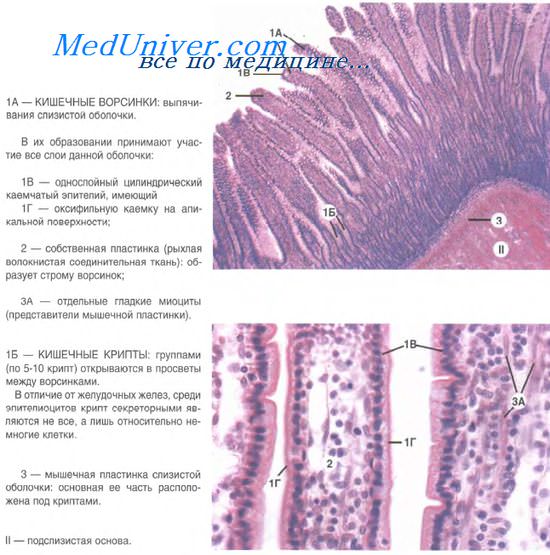
Слой рыхлой соединительной ткани практически всегда подстилает эпителиальный пласт. В слизистых оболочках она называется собственная пластинка слизистой. Здесь расположены капилляры, благодаря которым осуществляется питание эпителиальных клеток, поскольку в самом эпителии кровеносные сосуды отсутствуют. Поступление веществ идет по механизму диффузии (обязательно пересекая базальную мембрану), и именно эта особенность лимитирует толщину эпителиев. Наиболее удаленные от сосудов клетки погибают.
Общее строение и функции клетки
Клетка представляет собой наименьшую структурно-функциональную единицу живых организмов, способную к самообновлению, саморегуляции и самовоспроизведению.
Клетка — это специализированная система биополимеров, ограниченная клеточной мембраной, обладающей избирательной проницаемостью.
Все клетки подразделяют на прокариотические, не содержащие отдельное ядро, — примитивные одноклеточные организмы и эукариотические, в состав которых входит ядро с ядерной оболочкой, отделяющей генетическую информацию от цитоплазмы.
Надцарство эукариот включает в себя царства грибов, растений и животных. У эукариот биополимеры образуют ядро и цитоплазму, обеспечивая совокупность метаболических и энергетических процессов, осуществляя поддержание и воспроизведение всей системы в целом.
Клетки животных и растений характеризуются некоторыми специфическими особенностями. Растительная клетка окружена сложно устроенной клеточной оболочкой, содержит пластиды и некоторые другие структуры. Клетки животных состоят из ядра, цитоплазмы и цитолеммы (клеточной мембраны) и при всем структурном разнообразии имеют общие черты строения.
Клетка — это та наименьшая структура, которая обладает всеми основными свойствами живого. Ее части и органеллы не способны по отдельности выполнять все функции и, таким образом, не могут рассматриваться как отдельные (автономные) живые единицы. В то же время изучение частей клетки, особенно на субмолекулярном уровне, в течение последних 20…30 лет расширили представления о механизмах возникновения и развития врожденных и приобретенных болезней.
Несмотря на общую организацию, клетки могут быть весьма разнообразны по форме, а их строение зависит от положения и специализации. Однако при микроскопии чаще видно не саму клетку, а ее срез. В связи с этим приведены некоторые из возможных срезов клетки веретеновидной формы. О форме клетки можно судить при изучении ее в нескольких проекциях, дополнительной мысленной реконструкции объекта или используя специальные методы, позволяющие прямо получать объемное изображение микроструктуры — сканирующая электронная микроскопия.
Кроме клеток в организме имеются межклеточное вещество, неклеточные (симпласт, синцитий) и постклеточные структуры. Симпласт — это многоядерное образование с единой цитоплазмой и клеточной мембраной.
В синцитии клетки соединены между собой цитоплазматическими мостиками. Постклеточные структуры обычно являются частями клеток, например, тромбоцит млекопитающих — это часть цитоплазмы гигантской клетки — мегакариоцита. Все эти структуры — производные клеток.
Межклеточное вещество образовано гомогенным (основным, аморфным) веществом, или матриксом, и волокнами. В различных тканях соотношение волокон и аморфного вещества, а также их химический состав и распределение различны. Так, в рыхлой волокнистой соединительной ткани преобладает аморфное вещество, имеющее свойства сильно разведенного геля.
Коллагеновые и эластические волокна располагаются в виде сети. В сухожилии основной объем ткани занимают упорядоченно лежащие пучки коллагеновых волокон. В гиалиновом хряще развиты как волокна, так и аморфное вещество. Аморфное вещество в хряще имеет свойства плотного желе. В эпителиальной ткани межклеточное вещество слабо развито и основной объем занимают клетки.
Основные части (структуры) клетки — это цитоплазма и ядро. Каждая из этих структур жизненно необходима для клетки эукариот, имеет сложную биохимическую и макромолекулярную структуру и содержит свой набор ферментных систем.
Кроме того, клетка состоит из различных по составу и выполняемой функции, отделенных друг от друга мембранами компартментов (отсеков): гиалоплазмы, кариоплазмы, перинуклеарного пространства, гранулярной эндоплазматической сети (ЭПС), гладкой ЭПС, комплекса Гольджи, лизосом, пероксисом, митохондрий, включений, в том числе транспортных и секреторных пузырьков.
Жизнь в клетке возможна только при условии, что разнообразные ферменты и другие вещества упорядоченно распределены. Это обеспечивают мембраны. Они не дают смешиваться содержимому одного отсека с другим, так как там идут непрерывные биохимические реакции, пространственные перемещения. Мембранные структуры обособляют содержимое клетки от внешней среды или отделяют различные компартменты внутри клетки.
Компартментализация клетки обеспечивает межмолекулярные взаимодействия, наиболее эффективное течение биохимических процессов, возможность поляризации структур клетки.
Поляризация (полярность) клетки — это неравномерность в распределении органелл и других структур в клетке, концентрации ферментных комплексов клеточной мембраны, что сопровождается различиями в выполняемой функции. Поляризация встречается во многих специализированных клетках и неклеточных структурах: в эпителии, нервной ткани, остеокластах, ретикулоэпителиальных и ретикулоэндотелиальных клетках и др.
Важной особенностью мембран является возможность избирательной проницаемости. Проницаемость мембраны для того или иного вещества зависит от набора рецепторов и ферментов, имеющихся в мембране. Каждый из компартментов отличается также и по содержанию рецепторных белков мембран. Это обусловливает различия в проницаемости, выполняемых функциях, локализации и клетке, особенностях трансмембранного переноса веществ.
Компартменты сформированы в клетке в результате длительного эволюционного развития и передаются от одной клетки к другой при делении, как и генетический материал (по сути как внеядерная наследственность). Потеря одного из компартментов в процессе деления невосполнима и не может быть восстановлена за счет генома. Геном лишь привносит индивидуальные особенности к уже имеющимся структурам. К тому же часть компартментов (митохондрии животных, пластиды растений) содержат собственный генетический аппарат, значительно отличающийся от генома ядра, частично контролирующий свойства самих органелл.
Мембранные структуры клетки активно регулируют содержимое компартментов, контролируют физико-химические свойства, препятствуя диффузии либо, наоборот, — накапливая определенные молекулы.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Полярность и центросомы
Е. Каудри, «Раковые клетки»
Под ред. проф. В. В. Алпатова и др.,
Издательство иностранной литературы, М., 1958 г.
OCR Wincancer.Ru
Приведено с некоторыми сокращениями
Для цитолога полярность является столь же важной категорией, как и для физика. Клетка организована так, что она, подобно магниту, имеет два полюса. Многие компоненты клетки ориентированы между этими двумя противоположными полюсами. Мы остановимся на вопросе о полярности клетки как целого.
Эта полярность наиболее очевидна в тех случаях, когда условия у каждого полюса различны и функциональная активность клетки направлена в сторону одного из полюсов. Железистые клетки имеют проксимо-дистальную полярность: от проксимального, внутреннего полюса, расположенного вблизи тканевой жидкости, откуда клетка снабжается пищевыми веществами, к дистальному полюсу, откуда продукты секреции выделяются в проток железы, сообщающийся с внешней средой. В эпителиальных тканях, не имеющих выводных протоков, где клетки образуют пласты, располагающиеся один над другим, например в эпидермисе, проксимо-дистальная полярность выражена слабее, так как клетки в таких тканях не имеют удлиненной формы (т. е, не выгнуты в направлении переноса поступающих в клетку веществ). Нервные клетки поляризованы в направлении проведения нервного импульса. Как правило, в мезенхимных клетках полярность выражена менее определенно.
В эпителиальных клетках, обладающих проксимо-дистальной полярностью, ось клетки проходит через середину ядра и через центр клетки. Этот центр играет активную, даже доминирующую роль в процессах клеточного деления и поэтому имеет для нас большое значение. В неделящихся клетках в клеточном центре иногда удастся видеть центральную зону (микро-центр), чаще называемый центросомой. Внутри центросомы при благоприятных условиях можно видеть одну или две центриоли; в последнем случае центросома называется диплосомой.
Полярность меланоэпителиоматозных клеток доказывается тем, что центросомы располагаются в таких клетках между ядром и капиллярной сетью (Массон, 1933). В злокачественных клетках, инвазирующих ткани поодиночке или группами, полярность выражена слабо, значительно слабее, чем в эпителиальных клетках эмбрионов, которые ориентируются на очень ранних стадиях.
По данным Цвейбаума (1933), в культурах злокачественных макрофагов центросомы видны более отчетливо, чем в культурах нормальных макрофагов. Детального и непосредственного сравнения центросом злокачественных клеток и их нормальных прототипов на окрашенных срезах тканей до сих пор не проведено.
В раковых клетках, так же как и в нормальных (при некоторых особых условиях), число центросом может увеличиться. Борель (1901) и Ле-Конт (1902) показали, что в некоторых клетках карцином центросомы увеличиваются и образуют структуры, не отличимые от так называемых «телец Плиммера». Однако было бы неправильно делать вывод о том, что все тельца Плиммера и включения в виде «птичьего глаза» возникают из центросом.
На препаратах, окрашенных гематоксилин-эозином, центросомы обычно не видны. После соответствующей фиксации центросомы иногда четко выявляются при окраске железным гематоксилином. В живых клетках центросомы могут остаться незамеченными, но выявляются при фазово-контрастной микроскопии.
Поликар и Бессис (1953) провели детальное микрокинематографическое и электронно-микроскопическое исследование лейкоцитов человека и некоторых животных. При микрокиносъемке с применением фазового контраста центросомы обнаруживаются в виде области, лишенной видимых гранул и располагающейся в углублении ядра. Между лучами, идущими от центросомы, видны токи жидкости, направленные от периферической цитоплазмы к центросоме и обратно. Наибольший интерес представляют ритмические колебания центросом, имеющие амплитуду 3—7 m и частоту около 1 колебания в 1 мин. Направление колебаний параллельно длинной оси ядра и плоскости прикрепления клетки к поверхности, на которой она располагается. После каждого полупериода колебания наблюдается фаза покоя продолжительностью в несколько секунд. При понижении температуры от 37 до 4° амплитуда колебаний постепенно уменьшается; при 4° колебания прекращаются. Исследования электронно-микроскопических препаратов, приготовленных методом напыления, показали, что центросома представляет собой своего рода «холмик», доминирующий над «плоскогорьем» ядра и над еще более низким уровнем цитоплазмы. Это, вероятно, указывает на то, что центросома содержит меньше воды и больше белков (сухого вещества), чем ядро и цитоплазма. Эти исследования открывают путь для сравнительного изучения злокачественных лейкоцитов при помощи тех же методов.
 |
 Питание при ракеКаким должно быть питание при онкологических заболеваниях? Какие продукты абсолютно противопоказаны при той или иной форме рака?
|
 Узнать подробности >>
Узнать подробности >> |
 Фитотерапия в онкологииФитотерапия способна оказать существенную помощь не только в лечении онкологических заболеваний, но также и в их профилактике.
|
|
 Наследственность и ракМногих людей, имеющих у себя или у родственников онкологическое заболевание, интересует вопрос: передается ли рак по наследству?
|
|
 Рак при беременностиЛечение рака во время беременности является довольно сложным, ведь большинство лекарственных средств обладает токсичностью.
|
|
 Беременность после ракаКакие перспективы у беременности после перенесенного онкологического заболевания? Следует ли выдерживать срок после лечения рака?
|
|
 Профилактика ракаПрофилактика является важной частью общей борьбы с онкологическими заболеваниями. Как же уменьшить вероятность возникновения рака?
|
|
 Паллиативное лечение ракаЧто представляет из себя паллиативное лечение рака? Как оно может повлиять на качество жизни онкологического больного и изменить ее к лучшему?
|
|
 Новые методы лечения ракаУчеными разработано достаточно много перспективных методов лечения рака, пока еще не признанных официальной медициной. Но все может измениться!
|
|
 Статистика онкозаболеванийСтатистика заболеваемости раком, к сожалению, неутешительна: наблюдается рост числа заболевших, при этом болезнь «молодеет».
|
|
 О «народной» медицине
|
|
 Как бороться с раком?Как найти силы для борьбы с раком? Как не впасть в отчаяние от возможной инвалидности? Что может послужить надеждой и смыслом жизни?
|
|
 Как помочь близким?Как помочь близкому человеку жить с диагнозом «рак»? Нужна ли «ложь во спасение»? Как вести себя, чтобы близкие люди меньше страдали?
|
|
 Стресс и ракБытует такое мнение, что постоянные стрессовые ситуации способны привести к развитию онкологических заболеваний. Так ли это?
|
|
 Борьба с кахексиейМногие онкологические больные часто страдают от резкой потери веса. Чем это вызвано и можно ли как-то справиться с этой проблемой?
|
|
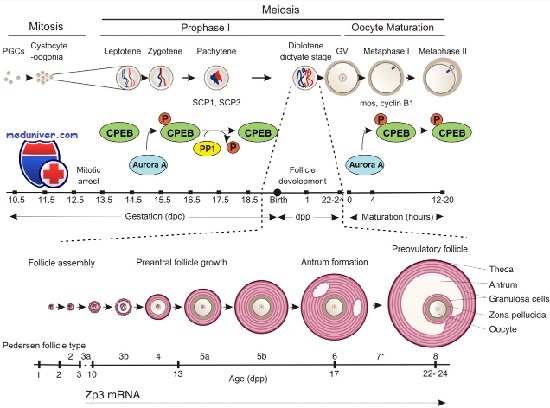
 Уход за лежачими больнымиПравила ухода за больными, вынужденными постоянно находиться в кровати, имеют свои особенности и их нужно обязательно знать. Полярность клетки обеспечивается чемВ отличие от других биологических моделей, например плодовой мушки, механизмы формирования полярности клетки, т.е. асимметричного распределения содержимого цитоплазмы, в ооците и эмбрионе млекопитающих до конца не ясны. Одна из сложностей заключается в том, что, хотя полярность, проявляющаяся в виде эксцентричного расположения ГП, наблюдается в ооцитах некоторых млекопитающих, например человека, резус-макаки, коровы и свиньи, ооциты стадии ГП, полученные из яичников крыс и мышей, такой полярности не демонстрируют, возможно, из-за артефактов, связанных с их культивированием in vitro. У растущего ооцита существует один главный центр организации микротрубочек (ЦОМТ). В состав ЦОМТ входит центросома, которая обеспечивает астральную конфигурацию микротрубочек. В клетках, делящихся путем митоза, центросомы образованы двумя центриолями, представляющими собой цилиндрические структуры, которые содержат девять триплетов микротрубочек, формирующих астральные волокна, необходимые для деления клетки. У центросом ооцита нет центриолей, поэтому они получили название ЦОМТ. В зрелом, мейотически компетентном ооците присутствуют множественные ЦОМТ, которые в процессе созревания ооцита обеспечивают миграцию герминативного пузырька (ГП) из центрального положения в эксцентрическую (кортикальную) позицию. Эксцентрическое положение ГП связано с положением мейотического веретена (или даже определяет его положение), которое в свою очередь определяет место отделения полярного тельца. Эксцентрическое положение мейотического веретена деления играет роль в формировании анимально-вегетативной оси. В последней анимальный полюс содержит ассоциированный с веретеном деления комплекс, а вегетативный — скопления эндоплазматического ретикулума. Предполагают, что асимметричная организация цитоплазмы чрезвычайно важна для того, чтобы при первом и втором делении мейоза образовывались полярные тельца, а не дочерние клетки одинаковых размеров. Спорным остается вопрос о том, предопределяет ли второе деление мейоза точку слияния сперматозоида и яйцеклетки, положение первого митотического веретена и даже пространственную организацию первого митотического деления. Mos/митогенактивируемая протеинкиназа-сигнальный путь, регулирующий задержку метафазы II, участвует также и в возврате мейотического веретена из кортикального положения в центральное, характерное для митотического веретена. В связи с вышесказанным, факторы, детерминирующие полярность ооцита, по-видимому, оказывают влияние на развитие и выживание раннего эмбриона, характер которого, к сожалению, пока не известен. Разумеется, цитоплазма претерпевает грандиозную реорганизацию: от асимметричного ооцита и яйцеклетки до эмбриона, способного к симметричным делениям. Полярность клетки обеспечивается чемДеятельность клеток скелетных мышц не зависит от полярности клеток. Например, мышечные клетки получают сигналы о поступлении глюкозы от любой точки цитоплазматической мембраны, и, насколько можно в настоящее время предполагать, изменения в экспрессии генов не зависят от места попадания глюкозы в клетку. Клетки в составе эпителия различаются по своей полярности. Это разделение между апикальной (граничащей с содержимым кишки в случае кишечного эпителия) и базолатеральной частями является существенным для активности эпителия. Данное свойство клеток хорошо изучено относительно переноса нутриентов через эпителий кишки. Существуют различия в переносе через апикальную и базолатеральную мембраны ионов, молекул малого размера и макромолекул. Именно полярность эпителия определяет направление движения этих субстанций через эпителий (внутрь клетки или наружу из нее). Данное свойство следует учитывать также и при обсуждении других функций эпителиальной клетки. Полярность особенно важна при исследовании взаимодействия нутриентов и генов в кишечном эпителии. Полярность эпителиальных клеток характеризует два важнейших механизма, посредством которых нутриенты (и другие компоненты содержимого кишки) оказывают влияние на гены: Более того, кишечный эпителий выступает в роли барьера между организмом и внешней средой, которую в данном случае представляет содержимое кишки. Этот барьер не является полностью непроницаемым, поскольку кишечник выборочно «позволяет» макромолекулам проходить через него. Кроме того, происходит активная абсорбция нутриентов. Постепенно стало понятно, что энтероцит выступает в роли иммунной клетки. Он имеет рецепторы для продуктов деятельности бактериальных клеток, а также участвует в экспрессии разнообразных молекул на своей поверхности, имеющей иммуноглобулиновые домены. Эпителиальная клетка также экспрессирует белки, которые могут взаимодействовать с иммуноцитами в кишечнике. К этим белкам относят такие поверхностные молекулы, как молекулы главного комплекса гистосовместимости (ГКГС) класса II и те цитокины, которые высвобождаются из эпителия, например хемокины или интерлейкин (IL) 6.
Перечисленные сигнальные белки дают возможность эпителиальной клетке играть ведущую роль в процессах, происходящих в кишечнике. Исследовательская группа выдвинула гипотезу, что изменения содержимого кишечника регулируют экспрессию эпителиальной клеткой сигнальных молекул. Это означает, что изменения содержимого кишечника в результате вариаций в диете оказывают влияние через эпителий кишечника на другие его структуры, особенно на иммунную систему его слизистой оболочки. Существуют два компонента сигнального пути, связывающие питание и бактерии, находящиеся в просвете кишки, с иммунной системой слизистой оболочки: Современная наука позволяет изучить изменения, происходящие с молекулами в эпителии кишки в ответ на изменения в диете (афферентный путь). В качестве экспериментальной модели может выступить экспрессия молекул ГКГС класса II в эпителии мыши, которая только что перестала получать грудное молоко. Молекулы ГКГС класса II ответственны за презентацию антигена, их экспрессия на эпителии кишечника мышей отмечается после их отлучения от груди, поэтому возможен перевод мышей на привычную для них пищу (корм для мышей) или определенного состава жидкую смесь (энтеральное питание) для изучения разницы между двумя видами питания. Эпителиальные клетки мыши исследовали в разные моменты времени после прекращения кормления ее грудным молоком. При этом изучали экспрессию молекул ГКГС класса II и инвариантную цепь, которая также подвергается экспрессии. Воздействие энтерального питания (Vivonex) значительно отличалось от такового при кормлении мышей обычным для них кормом. В норме корм для мышей индуцирует экспрессию генов сигнальных молекул между 20 и 30 днями жизни, в то время как энтеральное питание за этот же период не приводит к экспрессии генов в кишечном эпителии. В настоящее время доказано, что регуляторный нуклеарный белок (трансактиватор класса II, СIIТА) является необходимым и достаточным условием для экспрессии молекул ГКГС класса II.
Были проведены эксперименты с целью изучить, действительно ли влияние питания опосредовано трансактиватором класса II. У мышей были выявлены три изоформы СИТА. Интересно, что нормальная комплексная диета повышала экспрессию молекул ГКГС класса II посредством СIIТА IV. Кроме регуляции молекул ГКГС класса II посредством диеты отмечалась еще и медленная, зависимая от времени регуляция посредством СIIТАIII. Данные эксперименты показывают, что изменения в диете соответствуют определенным изменениям молекулярных взаимосвязей между содержимым кишечника и механизмом сигнальной трансдукции в эпителиальной клетке. Бактериальная ферментация тех углеводов, которые не абсорбировались в кишечнике, приводит к продукции короткоцепочечных жирных кислот (КЦЖК). Это означает, что уровень бутирата отражает изменения в популяции бактерий и в тех субстратах, которые доступны для метаболизма бактерий. Уровень бутирата сильно зависит от внешних воздействий. Например, у новорожденных детей отмечают очень низкие уровни бутирата как в тонкой, так и в толстой кишке; уровень бутирата в организме ребенка достигает уровня взрослого к 2-летнему возрасту. Интересно, что в течение первых 6 мес жизни уровни бутирата намного выше у детей, находящихся на искусственном вскармливании, чем у детей, получающих грудное молоко. Следовательно, уровень бутирата отражает те изменения, которые происходят в содержимом кишечника. Было высказано предположение, что концентрация бутирата может изменять активность сигнала в эпителиальной клетке. Исходя из этой гипотезы было исследовано воздействие бутирата на IL-8 и экспрессию моноцитарного хемотаксического протеина 1 (МСР-1). Увеличение концентрации бутирата повышало секрецию IL-8 при одновременном снижении экспрессии МСР-1. Эти эффекты были обнаружены в популяциях неактивных эпителиальных клеток, но наибольшие изменения были характерны для клеток, стимулированных такими провоспалительными агентами, как липополисахариды (ЛПС) или IL-1b. Известно, что бутират изменяет ацетилирование гистонов. Нуклеосома состоит из соленоида гистонов, вокруг которого дважды «обернута» ДНК. Бутират повышает ацетилирование гистона, тем самым снижая его компактность. В результате ДНК не может «обернуться» вокруг большой нуклеосомы полным оборотом, а нуклеосома теряет способность упаковываться в плотную структуру. Это «обнажает» ДНК и делает ее более подверженной действию факторов транскрипции. Было высказано предположение, что таким образом бутират изменяет экспрессию хемокинов. Для проверки данной гипотезы был использован фунгицид трихостатин A (TSA), в 700 раз более мощный индуктор ацетилирования гистонов по сравнению с бутиратом. Если бы действие бутирата на секрецию хемокина было обусловлено повышением ацетилирования гистонов, то можно было бы ожидать подобного эффекта и от TSA. Опыты с TSA показали, что TSA повышал секрецию IL-8 и снижал секрецию МСР-1. В нижней части рисунка продемонстрировано, что и TSA, и бутират повышали ацетилирование гистона-4. Неацетилированные гистоны быстрее продвигались через гель и образовывали одну полоску («ступеньку»), в то время как ацетилированные гистоны образовывали «лестницу» в зависимости от степени ацетилирования. Гистон-4 имеет 4 ацетилированных остатка лизина. Таким образом, ацетилирование гистонов будет отражено в образовании «лестницы» из 5 «ступенек». Это можно увидеть при действии бутирата или TSA. Верхняя часть рисунка показывает, что TSA дает такой же эффект, как и бутират. TSA повышает секрецию IL-8 и снижает секрецию МСР-1. Более того, степень увеличения IL-8 меняется в зависимости от степени ацетилирования гистонов. Эффект бутирата на ацетилирование гистонов обратим.
После прекращения действия бутирата ацетилирование гистонов возвращается к норме и отмечается такое же действие IL-1b на секрецию IL-8 и секрецию МСР-1, как до эксперимента. КЦЖК с различной длиной цепи оказывают дифференцированное влияние на ацетилирование гистонов. Бутират является наиболее действенной КЦЖК, индуцирующей ацетилирование гистонов, в то время как от кислот с большей или меньшей углеводной цепью наблюдаются менее выраженные эффекты. Данное влияние на ацетилирование гистонов обусловлено действием на экспрессию IL-8 и МСР-1. Из всех использованных в опыте КЦЖК именно бутират вызывает наибольшее увеличение секреции IL-8 и наиболее выраженное уменьшение секреции МСР-1. Итак, эти опыты показывают, что бутират изменяет экспрессию хемокинов в эпителиальной клетке. КЦЖК изменяют экспрессию посредством ацетилирования гистонов. Данные эксперименты, однако, не исключают возможность того, что дополнительное действие бутирата может быть опосредовано промоторными системами. Действительно, недавно проведенные исследования показали, что бутират снижает экспрессию связывающего инсулиноподобный фактор роста белка (IGFBP) 3, посредством ацетилирования протеина, ингибирующего связывание ДНК. В дальнейшем необходимо исследовать взаимосвязь между хромосомной регуляцией, продемонстрированной в приведенном эксперименте, и регуляцией, обусловленной промотором, как для бутирата, так и для других молекул содержимого кишечника. Доказательство действия экспрессии генов эпителиальных клеток на иммунную систему слизистой оболочки (эфферентного пути) основано на возможности селективного изменения экспрессии генов в эпителиальной клетке кишечника посредством трансгенных технологий. В качестве модели для демонстрации управления иммунной системой слизистой оболочки эпителием была использована экспрессия хемокинов в эпителии. Хемокин IL-8, который у человека отвечает за мобилизацию нейтрофилов, был первым из идентифицированных хемотаксических цитокинов. Однако IL-8 у мышей не экспрессирован. Для изучения действия хемокинов на иммунную систему слизистой оболочки была разработана система, с помощью которой макрофагальный воспалительный белок 2, имеющий эффект, практически подобный эффекту IL-8 у человека, был присоединен к промотору FABPI (белок кишечника, связывающий жирную кислоту). Промотор активен только в эпителиальных клетках тонкой кишки и проксимального отдела толстой кишки. Была разработана специальная «конструкция», где промотор FABPI и макрофагальный воспалительный белок 2 комплементарной ДНК (кДНК) были присоединены к интрону и полиадениловому «хвосту». Эта «конструкция» была внедрена в яйцеклетку мыши. Было выявлено, что эпителий первой генерации экспрессирует макрофагальный воспалительный белок 2 мРНК. Анализы показали воздействие этой «конструкции» на мобилизацию как нейтрофилов, так и лимфоцитов. У трансгенной мыши отмечали повышенную мобилизацию нейтрофилов в собственной пластинке слизистой оболочки кишечника и в эпителиальных клетках. Эффект хемокина был отмечен только в тех клетках, где промотор FABPI был активен. В тонкой кишке, где промотор FABPI активен, мобилизация нейтрофилов, выражающаяся активностью миелопероксидазы (на единицу массы кишки), была значительно выше у трансгенных мышей. В проксимальной кишке, где промотор FABPI также активен, отмечали увеличение нейтрофильной инфильтрации. Однако в дистальном отделе толстой кишки, где промотор FABPI неактивен, подобных эффектов отмечено не было. Кроме того, в печени и селезенке трансгенных мышей не выявляли увеличения инфильтрации больше, чем у нормальных мышей (в этих органах промотор FABPI неактивен). На первый взгляд, эти данные свидетельствуют о том, что эпителиальная клетка может посредством высвобождения хемокинов изменять иммунную функцию слизистой оболочки в кишечнике in vivo.
Дальнейший анализ иммунной системы показал, что в тонкой кишке помимо инфильтрации нейтрофилов была отмечена повышенная инфильтрация лимфоцитов. Их количество в собственной пластинке слизистой оболочки было увеличено. Также было отмечено удвоение числа лимфоцитов в эпителиальном слое слизистой оболочки кишки. Исследование рецепторов, расположенных на поверхности лимфоцитов, локализованных в эпителии, показало, что лимфоциты экспрессировали CXCR2, который является рецептором, отвечающим за трансдукцию активности макрофагального воспалительного белка 2. Таким образом, эксперименты показывают, что изменение экспрессии только одного хемокина в эпителии влияет на функцию как лимфоцитов, так и нейтрофилов. Однако изменения содержимого кишечника могут оказывать воздействие на многие хемокины. Кроме того, вероятен эффект воздействия на иммунную систему и других цитокинов. Становится понятно, что изменения экспрессии генов в эпителии оказывают большое влияние на остальные составляющие иммунной системы слизистой оболочки. Существует гипотеза, что эти сигнальные процессы важны не только для понимания физиологии здорового организма, но и для лечения заболеваний. Например, начальным этапом терапии детей с болезнью Крона в Великобритании является лечебное питание. Несмотря на то что известны многие механизмы, посредством которых энтеральное питание проявляет свои лечебные свойства, есть мнение, что еще одним методом может быть радикальная смена содержимого кишечника до такой степени, что изменятся сигналы, идущие от эпителия кишечника к иммунному компоненту слизистой оболочки, в результате снизится экспрессия воспаления при болезни Крона. Этим методом можно достигать уменьшения активности воспалительного процесса при данной патологии. Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021 — Вернуться в оглавление раздела «физиология человека» |
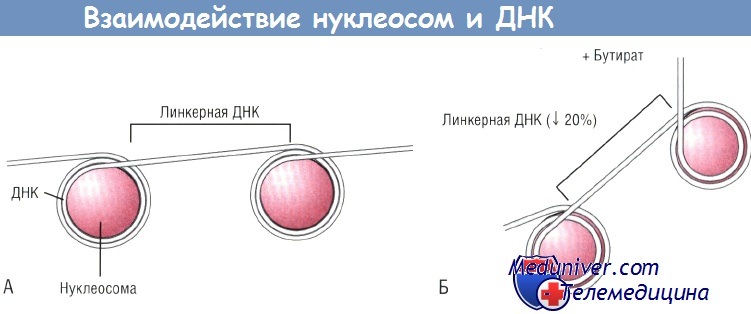 (А) ДНК делает два полных оборота вокруг нуклеосом, состоящих из неацетилированных гистонов.
(А) ДНК делает два полных оборота вокруг нуклеосом, состоящих из неацетилированных гистонов. 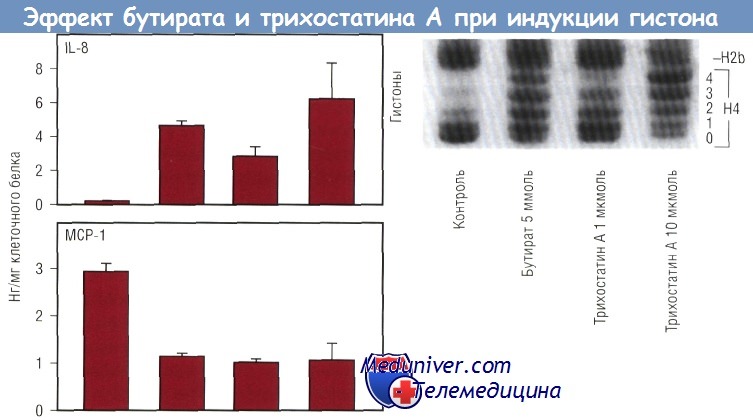 Ацетилирование и секреция хемокина клетками Сасо-2. Как бутират, так и трихостатин А увеличивали ацетилирование гистонов. Одновременно увеличивалась секреция IL-8 и уменьшалась продукция МСР-1 в клетках Сасо-2, стимулированных IL-1.
Ацетилирование и секреция хемокина клетками Сасо-2. Как бутират, так и трихостатин А увеличивали ацетилирование гистонов. Одновременно увеличивалась секреция IL-8 и уменьшалась продукция МСР-1 в клетках Сасо-2, стимулированных IL-1. 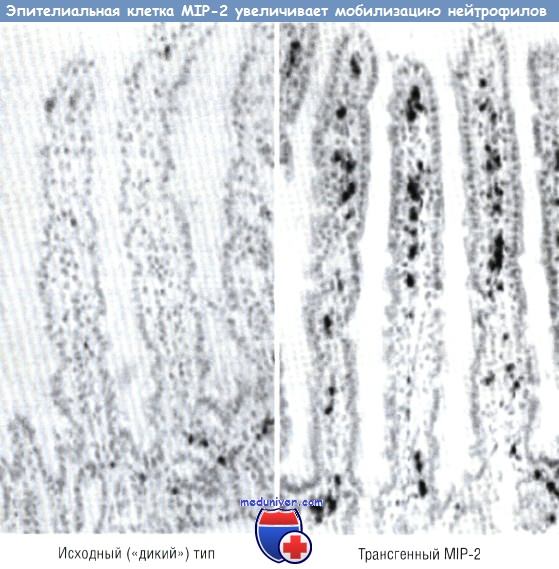 У трансгенной мыши MIP-2 экспрессирует эпителий кишечника, используя эпителий-специфический промотор. Нейтрофилы были локализованы с помощью миелопероксидазы.
У трансгенной мыши MIP-2 экспрессирует эпителий кишечника, используя эпителий-специфический промотор. Нейтрофилы были локализованы с помощью миелопероксидазы.