Почему учёные считают, что предками высших растений могли быть древние зелёные водоросли?
Почему учёные считают, что предками высших растений могли быть древние зелёные водоросли.

Главный признак представителей этого отдела отражён в названии.
Это самый многочисленный отдел водорослей, насчитывающий до 20 тыс.

К высшим споровым растениям относят : 1)мхи 2)бурые водоросли 3)лишайники 4)зелёные водоросли?
К высшим споровым растениям относят : 1)мхи 2)бурые водоросли 3)лишайники 4)зелёные водоросли.

Почему водоросли считают самыми древними растениями на Земле?
Почему водоросли считают самыми древними растениями на Земле?

Почему водоросли относятся к низшим растениям?
Почему водоросли относятся к низшим растениям?
Где обитают зелёные одноклеточные водоросли?
Какое строение имеет хламидоманада?
Среди высших растений обособленной и тупиковой ветвью развития принято считать : а)водоросли б)папоротниковидные растения в)моховидные растения г)голосеменные растения?
Среди высших растений обособленной и тупиковой ветвью развития принято считать : а)водоросли б)папоротниковидные растения в)моховидные растения г)голосеменные растения.

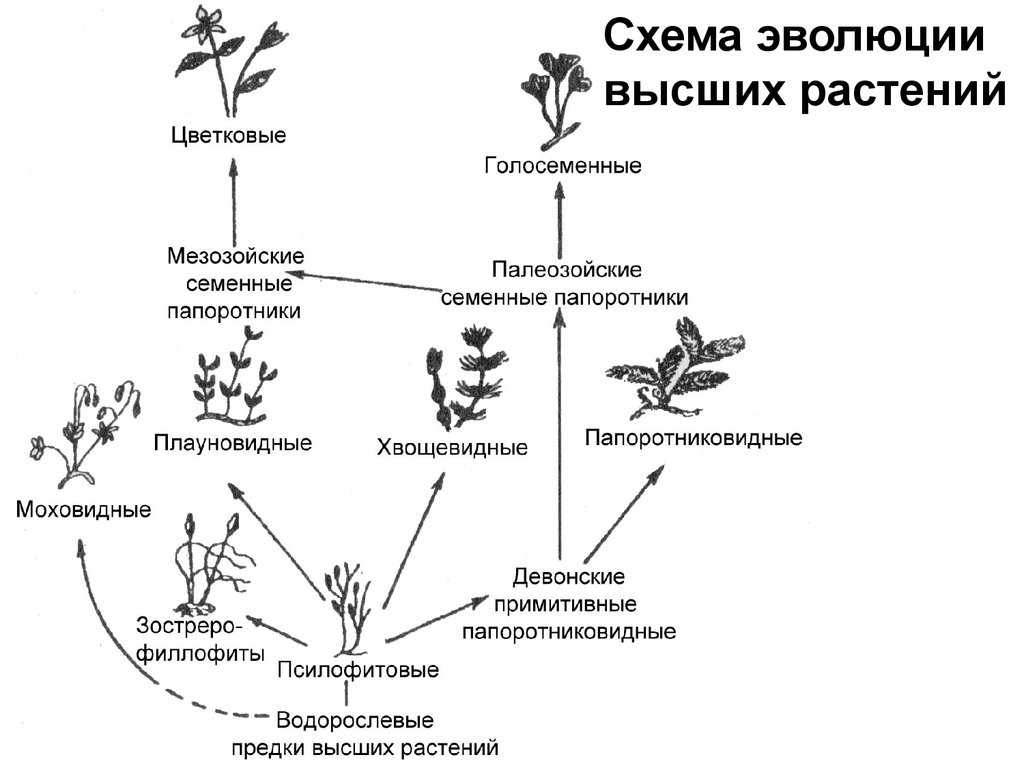
Составьте схему происхождение и эволюции высших растений используя следующие термины водоросли вы предки высших растений моховидных древние папоротники папоротники покрытосеменные плауновидные папорот?
Составьте схему происхождение и эволюции высших растений используя следующие термины водоросли вы предки высших растений моховидных древние папоротники папоротники покрытосеменные плауновидные папоротниковидные.

Водоросли считают просто организованными растениями почему?
Водоросли считают просто организованными растениями почему.
Чем различается водообеспеченность у водорослей и высших растений?
Чем различается водообеспеченность у водорослей и высших растений?
Водоросли это Высшие растения или нет?
Водоросли это Высшие растения или нет.
Чем схожи и чем различаются водоросли и высшие растения?
Чем схожи и чем различаются водоросли и высшие растения?



1)эксперимент Условия контролируются кем то значит они проводят эксперимент.

Признаки, свидетельствующие о древнем происхождении кишечнополостных : Полости тела отсутствует. Клетки слабодифференцированы, не образуют ткани и органы. Присутствуют только два зародышевых листка : эктодерма и энтодерма. Лучевая (радиальная) сим..
Если после употребления в пищу грибов появились признаки отравления, то необходимо срочно вызвать врача. До его прихода рекомендуется постоянное употребление жидкости. Крепкий холодный чай, а так же воду.
Возможные предки высших растений. Время и условия возникновения высших растений
Содержание:
| Предмет: | Ботаника |
| Тип работы: | Реферат |
| Язык: | Русский |
| Дата добавления: | 03.07.2019 |
Если вам тяжело разобраться в данной теме напишите мне в whatsapp разберём вашу тему, согласуем сроки и я вам помогу!
По этой ссылке вы сможете найти рефераты по ботанике на любые темы и посмотреть как они написаны:
Посмотрите похожие темы возможно они вам могут быть полезны:
Введение:
Высшие растения, или Embryobionta, – зародышевые растения, или Cormophyta, Cormobionta – побеговые растения, или Telomophyta, Telomobionta – теломные растения. Это одно из двух подцарств растительного мира. Высшие растения объединяют свыше 300 тыс. видов следующих таксономических групп (отделов): риниевые (псилофиты), моховидные (печёночники и мхи), псилотовые (псилотум и тмезиптерис), плауны, хвощи, папоротники, голосеменные и цветковые, или покрытосеменные.
В отличие от низших растений высшие представляют собой более сложные, дифференцированные многоклеточные организмы, которые приспособлены к жизни на суше (за исключением немногочисленных и явно вторичных водных форм), имеют правильное чередование двух поколений, т. е. полового (гаметофит) и бесполого (спорофит). На спорофитах у них развиваются многоклеточные спорангии, наполненные имеющими прочную оболочку неподвижными спорами, на гаметофитах развиваются многоклеточные половые органы (гаметангии). Однако у некоторых голосеменных (часть гнетовых) и у всех цветковых растений гаметангии в процессе эволюции исчезли.
Высшие растения как таксономическая категория
Высшие растения имеют таксономический статус. Они в ранге подцарства высших (Embryobionta), наряду с низшими (Thallobionta), относятся к царству растений (Vegetabilia, Chlorobionta, Plantae), надцарству эукариот (Eucaryota). Таким образом, их объединяют признаки таксонов ранга царства и надцарства. Как у эукариот, у высших и у низших растений присутствует оформленное ядро, снабжённое типичной оболочкой (двойная мембрана) и ядрышком; есть внеядерный митотический аппарат в виде микротрубочек, митохондрии, аппарат Гольджи, кинетосомы, жгутики и другие органеллы; клеточная стенка включает хитин или целлюлозу; представлены половой процесс и смена ядерных фаз, гаплоидной и диплоидной.
Высших растений в настоящее время, по-видимому, значительно больше 300 тыс. видов. И они распределяются по основным отделам следующим образом:
В отличие от настоящего корневища (модифицированного побега), корневище является первичным элементарным органом. Появление телома увеличило пластичность и открыло более широкие возможности для дальнейшего эволюционного развития. Конечное положение спорангиев, характерное для телом, у ринофитов приводит к тому, что спорангии представляют собой модифицированные верхушки ветвей.
Время появления высших растений
По имеющимся палеонтологическим данным, это произошло около 440 миллионов лет назад (силурский период палеозойской эры). Известна точка зрения, что некоторые водоросли, живущие недалеко от побережья, сначала «высунули головы в воздух», затем обосновались в зоне приливов, а затем, постепенно превращаясь в высшие растения, полностью сошли на берег. Впоследствии они постепенно завоевывали землю. Однако позднее было высказано серьезное палеонтологическое возражение против этой точки зрения.
Если процесс превращения водорослей в высшие растения действительно происходил в прибрежных водах, то в этом случае условия попадания в ископаемые записи самых старых высших растений наиболее благоприятны. Однако при обилии палеоботанического материала мы не видим ни одной из его «промежуточных» стадий.
Гаметангии из примитивных, часто одноклеточных, характерных для подавляющего большинства водорослей, превращаются в многоклеточные; их стены надежно защищают развивающиеся гаметы от неблагоприятных условий. В жизненном цикле первых наземных растений наблюдается регулярное чередование половых и бесполых поколений, как и в случае с их предками.
Мужские гаметы, сперматозоиды, развиваются на ростках антеридий, которые в сырую погоду или во влажных условиях, плавающие в воде, достигают архегония и сливаются с яйцом. Из-за небольшого размера гаметофитов оплодотворение у хвощей, хищений и папоротников может происходить даже в присутствии незначительного количества воды, такого как капли росы, туман.
К концу силурийского развития жизнь на Земле была отмечена значительным успехом. Так, в древних морях и океанах, полностью населенных к тому времени, все основные виды преимущественно водных животных, включая рыб, уже были представлены. Плотное население всех благоприятных мест обитания, интенсивная конкуренция между видами и внутри них, феномен «чрезмерной» рождаемости привели к необходимости развития новых мест обитания.
Гипотеза о происхождении высших растений
Известны две группы гипотез, которые рассматривают наследственные корни высших растений: альгологическую и симбиогенетическую. Согласно «водорослевым» гипотезам, образование высших растений происходит из какой-то группы водорослей. Коричневые, красные и зеленые водоросли были предложены на роль предков. У представителей первых двух отделов развиты половые органы, наиболее напоминающие таковые у высших растений. Однако, как доказали эмбриологи, это сходство является внешним, не важным для понимания природы археонии и anteridium. Их различия в физиологии, биохимии и цитологии особенно поразительны.
В то же время зеленые водоросли очень похожи на высшие растения по составу пигментов, особенностям клеточного деления, но отличаются очень примитивными гаметангами. Так, «зеленые» водоросли и высшие растения имеют «стандартный» набор пигментов: хлорофиллы А и В, астаксантин, лютеин, виолаксантин, неоксантин, зеаксантин.
Другими признаками являются наличие крахмала как основного продукта ассимиляции и клетчатки в клеточных стенках; осуществление синтеза лизина через диаминопимелиновую кислоту; двухчленные пластиды; центробежный цитокинез; расположение тилакоидов в хлоропластах; сплющенные или пластинчатые митохондриальные кристы; отсутствие фаготрофии.
Высшие растения в основном живут на суше, то есть в принципиально иных условиях по сравнению с водорослями (низшие растения). Когда первые растения пришли на сушу (примерно 400 миллионов лет назад), они были немедленно вынуждены адаптироваться к совершенно другим экологическим режимам. Все существенные факторы окружающей среды для растений на суше значительно различаются, чем в водной среде.
На суше растение теряет поддержку водной среды для своих надземных частей. В результате образуются механические ткани, которые противодействуют давлению ветра, дождя и снега.
Заключение
Общими чертами всех высших растений являются:
Присылайте задания в любое время дня и ночи в ➔ 

Официальный сайт Брильёновой Натальи Валерьевны преподавателя кафедры информатики и электроники Екатеринбургского государственного института.
Все авторские права на размещённые материалы сохранены за правообладателями этих материалов. Любое коммерческое и/или иное использование кроме предварительного ознакомления материалов сайта natalibrilenova.ru запрещено. Публикация и распространение размещённых материалов не преследует за собой коммерческой и/или любой другой выгоды.
ГДЗ биология 5 класс Пасечник, Суматохин, Калинова Просвещение 2019-2020 Задание: § Стр 61
Моя лаборатория
Строение зелёных водорослей
Помещаем на предметное стекло микроскопа каплю цветущей воды, накрываем покровным стеклом.
Рассматриваем при малом увеличении одноклеточные водоросли.
Оттягиваем часть воды из-под покровного стекла полоской фильтровальной бумаги и рассматриваем клетку водоросли при большом увеличении.
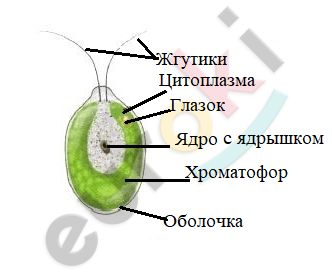
Водоросль состоит из одной клетки, а потому относится к низшим растениям. Форма тела округлая, немного вытянутая, обтекаемая. Есть два жгутика для передвижения.
Находим в клетке водоросли оболочку, цитоплазму, ядро, хроматофор. Обращаем внимание на форму и окраску хроматофора.
Клетка покрыта целлюлозной оболочкой. Хлорофилл, который нужен клетке для фотосинтеза, содержится в хроматофоре чашевидной формы. Есть ядро, цитоплазма, ядро с ядрышком, крахмальное тельце и пульсирующие вакуоли с клеточным соком.
Зарисовываем клетку и подписываем названия её частей.
Стр. 61. Вопросы после параграфа
№ 1. Где обитают зелёные одноклеточные водоросли? Какое строение имеет хламидомонада?
Большинство зеленых водорослей, в отличие от бурых или красных, обитает в пресноводных водоемах, а лишь некоторые их виды в морях. Также есть отдельные представители, которые приспособились к жизни и на суше, например, в почве и в сырых затемненных местах, на поверхности камней и зданий, на коре деревьев и валунах. В период засухи те виды, которые обитают в воде, находятся в состоянии покоя до наступления благоприятных условий.
В клетке хламидомонады расположен только один хлоропласт, в котором не только протекает процесс фотосинтеза, но и запасается крахмал. Хлорофилл, содержащийся в нем, придает хламидомонаде зелёную окраску.
№ 4. Где обитают и какое строение имеют зелёные многоклеточные водоросли?
Многоклеточные водоросли обитают в озерах, морях, стоячих водах и реках. Их тело или слоевище может иметь листовидную или нитевидную форму. В зависимости от пигмента различают зеленые, бурые и красные многоклеточные водоросли. Не редко в стоячих водах можно увидеть ярко-зеленого цвета комочки. Они состоят из скользких нитей зеленой многоклеточной водоросли спирогиры, клетки которой вытянутые в дину, имеют целлюлозную клеточную стенку и покрыты слизью. Между собой клетки соединены торцами и образую нити. Внутри у них располагается цитоплазма с закрученными в спирали лентовидными хроматофорами.
Еще одни представитель многоклеточных зеленых водорослей – улотрикс, чьи ярко-зелёные нити крепятся к различным подводным предметам. Нити этой водоросли образованы короткими и соединенными между собой клетками. В каждой из клеток есть ядро и хроматофор, который по форме напоминает незамкнутое кольцо. Растут нити улотрикса за счет деления его клеток.
В солёных водах морей и океанов можно обнаружить морской салат или ульву – морскую зеленую водоросль, тело которой пластинчатое, состоит только из двух клеток, а в длину достигает от 30 см до 1,5 метров.
Самым сложным строением отличаются пресноводные харовые водоросли, которые по внешнему виду напоминают хвощи. Один из представителей харовых водорослей – блестянка гибкая или нителла – используется в аквариумах. Ее слоевище чем-то напоминает строение высших растений, так как к грунту оно крепится при помощи нитевидных неокрашенных выростов – ризоидов, а части его могут выглядеть как листья и стебли.
№ 3. Что такое ризоиды?
Ризоиды представляют собой нитевидные выросты в нижней части «черешка» у водорослей, плаунов, хвощей, лишайников, мхов, которые нужны для прикрепления их слоевища к разным поверхностям и извлечения из них питательных веществ (из почвы, субстрата). В отличие от корней, ризоиды имеют более сложную структуру, потому что состоят из большого количества элементов.
№ 4. Где обитают и какое строение имеют красные водоросли?
Красные водоросли или, как их еще называют, багрянки обитают преимущественно в морях и, в отличие от бурых и зеленых водорослей, на больших глубинах. Только 200 видов из 4000 багрянок обитает в почвах и пресноводных водоемах. Кроме того, они могут обитать не только на глубине 100 – 200 метров, а отдельные их представители и на глубине 300 – 500 метров, но и в верхних горизонтах моря.
Красные водоросли, как правило, довольно крупные растения. Но встречаются среди них и микроскопические виды. Есть среди них и одноклеточные (очень редко), нитчатые и псевдопаренхимные формы. Клеточная стенка красных водорослей состоит из фибриллярного матрикса и аморфной фракции, в состав которой входят агароиды, агар, маннаны и каррагинаны. У некоторых представителей клеточная стенка инкрустируется карбонатами кальция, стронция и магния. А иногда поверх такой стенки располагается и белковая кутикула.
Запасные вещества в виде багрянкового крахмала запасаются в цитоплазме. Жгутиковые стадии жизненного цикла полностью отсутствуют.
Хлоропласты красных водорослей двумембранные и с одиночными тилакоидами, один или два из которых обычно лежат на периферии хлоропласта. Также на мембранах тилакоидов есть и фикобилисомы.
Основной пигмент хлоропластов у красных водорослей это хлорофилл. Но также у них есть каротиноиды и фикобилины, благодаря чему водоросли могут поглощать свет практически всей видимой частью спектра. В соответствии с изменениями соотношения фотосинтетических пигментов окраска их может меняться с увеличением глубины. Например, на мелководье они желто-зеленые или с голубым оттенком, потом становятся розовыми и на глубине от 50 метров приобретают насыщенный красный цвет.
№ 5. Где обитают и какое строение имеют бурые водоросли?
Бурые водоросли представляют собой отдел истинных многоклеточных водорослей, которые имеют соответствующую бурую расцветку. К этой группе относится около 1500 видов и 250 родов растений, в большинстве своем являющихся пресноводными формами. Распространены бурые водоросли в морях земного шара повсеместно, достигая многочисленности и разнообразия в холодных водоёмах умеренных и приполярных широт, где и формируют в прибрежных полосах большие заросли.
Есть бурые водоросли и в тропическом поясе. Их крупное скопление отмечается в Саргассовом море, а массовое развитие их происходит обычно зимой, когда температура воды понижена.
По размерам бурые водоросли могут быть от нескольких сантиметров и до нескольких десятков метров. Их многоклеточное слоевище окрашено от темно-бурого до оливково-зеленого цвета, потому что кроме хлорофилла в его клетках содержится еще и немалое количество желтых и коричневых пигментов. Из всех водорослей именно они отличаются наиболее сложным строением: у некоторых представителей клетки сгруппированы в один или два ряда, чем напоминают строение тканей высших растений.
Слоевище у бурых водорослей может быть разной формы: вертикально висящие или стелющиеся нити, ветвящиеся кусты или пластинки. Крепятся они к твердому субстрату при помощи ризоидов. Для высших бурых водорослей порядка фукусовых и ламинариевых характерной является не только дифференциация тканевых структур, но и появление проводящих систем. В отличие от представителей других групп, для них свойственно наличие и многоклеточных волосков с базальной зоной роста.
У каждой клетки в строении бурых водорослей покровом служит толстая клеточная стенка, которая состоит из 2 – 3 слоев, сильно покрытая слизью. Структурный компонент клеточной стенки – пектин и целлюлоза. В каждой клетке содержится только одно ядро и вакуоли, которых может быть как одна, так и несколько. Хлоропласты в клетках мелкие, дисковидной формы, имеют бурую окраску благодаря содержанию помимо каротина и хлорофилла ксантофиллов. Также в цитоплазме клеток откладываются и запасы питательных веществ: разные масла, многоатомный спирт маннит, полисахарид ламинарина.
Стр. 61. Подумайте
Почему учёные считают, что предками высших растений могли быть древние зелёные водоросли?
По мнению ученых, именно древние зеленые водоросли при наступлении благоприятных условий на суше смогли «выйти» в новую для них среду обитания, приспособиться к ней и стать началом в эволюционной ветке развития высших растений. Одной из причин связи зеленых водорослей и высших растений является наличие в составе цианобактерий пигмента хлорофилла в хроматофорах, который и обеспечивал им зеленую окраску, а также способность к фотосинтезу.
15. Роль водорослей в природе и жизни человека
Сценарий происхождения высших растений во многом похож на сценарий происхождения животных


Рис. 1. Некоторые стрептофиты. 1 — истинно многоклеточная (с дифференциацией органов и тканей) харовая водоросль Chara (фото Sven Dahlke с сайта dbg-phykologie.de). 2, 3 — пластинчатая колеохетовая водоросль Coleochaete (иллюстрации из статьи L. E. Graham et al., 2012. Aeroterrestrial Coleochaete (Streptophyta, Coleochaetales) models early plant adaptation to land): 2 — обычная жизненная форма водной колеохетовой водоросли: однослойная дисковидная пластинка (стрелочки указывают на клетки, имеющие хлоропласты особой формы), 3 — колеохетовая водоросль, выращенная в эксперименте вне воды: это уже не однослойная пластинка, а толстый «коврик» из переплетающихся нитей; длина масштабного отрезка на фото 2 — 50 мкм, а на фото 3 — 500 мкм = 0,5 мм (в десять раз больше!). 4 — нитчатая сцеплянковая водоросль Spirogyra (фото с сайта en.wikipedia.org). 5 — одноклеточная сцеплянковая водоросль Closterium (фото с сайта protist.i.hosei.ac.jp). 6 — спорофиты мха Rhizomnium (фото Margaret Gaines с сайта fineartamerica.com). Спорофиты — это коробочки на высоких ножках; «листочки» внизу — гаметофиты. Именно многоклеточный спорофит стал главным приобретением высших растений, отличающим их от всех других стрептофит
Исследования геномов высших растений и их родственников, относящихся к стрептофитовым водорослям, приводят биологов к двум выводам: 1) высшие растения не выходили на сушу, будучи многоклеточными, а изначально формировались прямо на суше — весьма вероятно, что их ближайшие одноклеточные предки уже были наземными; 2) молекулярные преадаптации к освоению суши формировались у предков высших растений очень постепенно и давно — как минимум с первых шагов эволюции ветви стрептофит, и в этом отношении история происхождения высших растений схожа с историей происхождения многоклеточных животных.
Зеленые хозяева Земли
Трудно найти событие, которое бы изменило облик планеты Земля сильнее, чем возникновение высших растений. Образованный ими сплошной растительный покров, который с середины палеозоя покрывает большую часть земной суши, можно было бы назвать одним из биологических чудес света; недаром изучением его структуры занимается отдельная сложная наука — геоботаника. Высшие растения сильно изменили состав земной атмосферы, обогатив ее кислородом в количестве, немыслимом в более ранние эпохи. Повлияли они и на климат, и на рельеф, и уж тем более — на круговорот биогенных элементов. Ну и само собой разумеется, что появление наземных растительных сообществ создало много новых экологических ниш с новыми источниками питания, а это стало «приглашением» к выходу на сушу многоклеточных животных — сначала членистоногих, потом и позвоночных.
По всем этим причинам неудивительно, что происхождение высших растений очень интересует биологов. Откуда же они взялись?
Высшие растения относятся к большой эволюционной ветви эукариот, которая называется Plantae или Archaeplastida (см. рис. 2 в новости Случайно открытый жгутиконосец обновляет систему эукариот, «Элементы», 06.02.2019). Если представить себе родословное древо эукариот в виде обычного дерева, то от основания ствола архепластид первым делом отойдут две не слишком толстые веточки: глаукофитовые водоросли и красные водоросли. Все остальные архепластиды образуют мощную ветвь, которая называется Viridiplantae, то есть — буквально — зеленые растения. Однако ветвь зеленых растений, в свою очередь, сразу разделяется на две ветви потоньше: хлорофиты и стрептофиты (см. Д. Леонтьев, 2014. Общая биология: система органического мира). Хлорофиты гораздо разнообразнее, но зато стрептофиты успешнее освоили сушу. Высшие растения — это веточка внутри группы стрептофит (рис. 1). Таково их «место прописки» на всеобщем эволюционном древе.
Надо сказать, что хлорофиты и стрептофиты во многом похожи друг на друга. Обе ветви начались с одноклеточных, и в обеих постепенно появилась многоклеточность, причем это происходило отчасти параллельно: без помощи микроскопа неспециалист вряд ли отличит, например, хлорофитовую нитчатую водоросль улотрикс от стрептофитовой нитчатой водоросли спирогиры, хотя общие предки этих водорослей, несомненно, были одноклеточными. Возникает вопрос: почему эти ветви вообще разошлись?
Немецкий биолог Буркхард Беккер (Burkhard Becker) обратил внимание на то, что между хлорофитами и стрептофитами есть различия в механизме фотодыхания (B. Becker, 2012. Snow ball earth and the split of Streptophyta and Chlorophyta; см. также обзор в журнале «Знание — сила»). Фотодыхание — это частичное «сжигание» продуктов фотосинтеза, сопровождаемое поглощением кислорода, выделением углекислого газа и синтезом аминокислот. В одном из этапов фотодыхания могут для одной и той же цели использоваться разные ферменты: у хлорофит — гликолатдегидрогеназа, а у стрептофит — гликолатоксидаза. Разница в том, что в первом случае побочным продуктом этого этапа становится молекула с замечательным названием «восстановленный никотинамидадениндинуклеотид» (НАДH), которая заодно служит источником энергии, а во втором — абсолютно ненужная клетке перекись водорода. То есть биохимия стрептофит менее экономична. Между тем вероятное время расхождения хлорофит и стрептофит, судя по молекулярным данным, лежит где-то в окрестности знаменитой эпохи Земли-снежка, когда большая часть земной поверхности, включая и сушу, и океаны, была покрыта толстым слоем льда (см. С. Ястребов, 2016. Кислородная революция и Земля-снежок). Беккер предполагает, что хлорофиты сформировались в океане под ледяным панцирем, где света для фотосинтеза было мало и приходилось бороться за каждый квант энергии, а стрептофиты — в пресных лужах на поверхности ледяного панциря, где могло не хватать всего остального, но уж свет был в избытке. Поэтому там, где была возможность выбора, хлорофиты предпочли более сложный и экономичный путь, а стрептофиты — более прямолинейный и расточительный. Если эта красивая гипотеза верна, то получается, что стрептофитовые водоросли уже с момента своего возникновения были «слегка наземными».
Однако для того, чтобы разобраться в проблеме происхождения высших растений, этого все равно недостаточно. Тут нужны детали, и прежде всего — касающиеся родственных связей.
Неожиданные родичи
Помимо высших растений, к стрептофитам относятся шесть групп водорослей: мезостигмовые, хлорокибовые, клебсормидиевые, харовые, колеохетовые и сцеплянковые. Первые три группы по всем параметрам относительно далеки от высших растений, и о них сейчас можно больше не говорить, к счастью для тех читателей, которых пугает альгологическая номенклатура. На протяжении многих десятилетий ближайшими родственниками высших растений считались харовые водоросли. Это — крупные многоклеточные пресноводные водоросли, внешне похожие на хвощ, с отлично выраженными аналогами стебля и листьев и с многоклеточными половыми органами, напоминающими половые органы высших растений. Более роскошных предков было не найти.
Существовала, впрочем, и альтернативная идея, согласно которой высшие растения произошли не от харовых, а от колеохетовых водорослей. Последние не достигают такой сложности, как харовые, но зато прекрасно приспосабливаются к жизни на мелководье. «Тело» такой водоросли представляет собой пластинку из переплетенных нитей, которую очень легко представить себе в качестве переходной стадии при выходе на сушу. Более того, в эксперименте колеохетовую водоросль можно заставить жить вообще вне воды, просто на влажном субстрате (L. E. Graham et al., 2012. Aeroterrestrial Coleochaete (Streptophyta, Coleochaetales) models early plant adaptation to land).
Однако похоже на то, что молекулярная филогенетика опровергает все эти гипотезы. На самом деле ближайшими современными родственниками высших растений оказались не харовые и не колеохетовые, а сцеплянковые водоросли (M. N. Puttick et al., 2018. The interrelationships of land plants and the nature of the ancestral embryophyte). Любой ботаник подтвердит, что из более-менее правдоподобных кандидатур именно сцеплянковые водоросли меньше всего похожи на высшие растения. В этой группе нет форм сложнее нитчатых (например, та же спирогира), большинство же сцеплянок — одноклеточные. Но самым серьезным доводом против близкого родства сцеплянок с высшими растениями всегда было размножение.
Дело в том, что у высших растений наблюдается типичная оогамия: маленькая подвижная мужская половая клетка оплодотворяет большую неподвижную женскую. При этом мужские половые клетки — сперматозоиды — как и у животных, двигаются с помощью жгутиков. У споровых растений, например у мхов и папоротников, жгутиковые сперматозоиды самостоятельно доплывают до яйцеклеток. Семенным растениям подвижность половых клеток не нужна, и у подавляющего большинства из них она вместе со жгутиками исчезла. Но интересно, что у двух групп семенных растений, а именно у саговников и у гинкго, жгутиковые сперматозоиды до сих пор есть (W. F. Friedman, E. M. Gifford, 1997. Development of the male gametophyte of Ginkgo biloba: a window into the reproductive biology of early seed plants). Это очень древний признак, остаток далекого эволюционного прошлого.
А вот у сцеплянок жгутиков не бывает: у них просто утрачены соответствующие гены. Поэтому их клетки всегда неподвижны. Одноклеточные сцеплянки придают себе плавучесть не за счет активного движения, а за счет причудливых выростов жесткой клеточной стенки. Надо сказать, что иногда это выглядит очень красиво: все желающие могут полюбоваться, например, на изображения относящихся к сцеплянкам десмидиевых водорослей, сделанные великим Эрнстом Геккелем. Но никаких сперматозоидов и яйцеклеток у сцеплянок нет. У них выработан оригинальный вариант полового процесса, когда клетки конъюгируют («сцепляются») и содержимое одной из них перетекает в другую. Ничего общего с размножением высших растений это не имеет.
Тем не менее, по современным данным, именно сцеплянковые водоросли — сестринская группа высших растений (рис. 2), точно так же, как воротничковые жгутиконосцы — сестринская группа животных (см. «Первичность губок» опережает по очкам «первичность гребневиков», «Элементы», 12.11.2019). Правда, эти жгутиконосцы все-таки не являются прямыми предками животных. Сцеплянки — тоже не прямые предки высших растений. Потеря жгутиков, изобретение особого способа размножения — все это наверняка случилось уже после того, как эволюционная ветвь сцеплянок отделилась от ветви, ведущей к высшим растениям. Но более близких современных родственников у высших растений, судя по всему, нет.

Рис. 2. Родственные связи стрептофит. Использована иллюстрация из статьи F. Leliaert et al., 2012. Phylogeny and molecular evolution of the green algae, но схема родственных отношений соответствует изложенной в обсуждаемой статье в Current Biology. Хлорокибовые водоросли не показаны: они не всегда считаются самостоятельной группой. Все остальные пояснения — в тексте
Что касается последнего общего предка высших растений и сцеплянок, то весьма вероятно, что он был одноклеточным. Ведь и сейчас большинство сцеплянок — одноклеточные. Это означает, что высшие растения при всем желании не могли унаследовать от своих водных предков никакие многоклеточные структуры. Наука, изучающая превращения таких структур — эволюционная морфология — тут бессильна. То ли дело, если бы предками высших растений оказались великолепные харовые. Но увы, это не так. Переход от водорослей к высшим растениям придется реконструировать «вслепую», без опоры на морфологическую преемственность.
Для ученых, работавших сто или хотя бы тридцать лет назад, такая задача была бы безнадежной. Она и сейчас непроста, но сейчас у биологов есть как минимум один новый мощный инструмент, позволяющий добыть хоть какую-то информацию. Это — чтение геномов.
Из воды на сушу. или наоборот?
В конце 2019 года вышла статья с анализом полных геномов двух сцеплянковых водорослей, находящихся очень близко к эволюционному корню этой группы (S. Cheng et al., 2019. Genomes of subaerial Zygnematophyceae provide insights into land plant evolution). Они называются Mesotaenium и Spiroglaea. Одна из них — спироглея — даже выделена авторами в особый подкласс: на филогенетическом древе, которое они получили, она оказалась сестринской группой ко всем остальным сцеплянкам, вместе взятым. Попутно проводились сравнения геномов сцеплянок с геномами других Viridiplantae: хлорофит, клебсормидиевых, харовых и высших растений. Серия таких сравнений позволяет довольно точно выяснить, какие гены когда приобретались.
В результате был составлен список генов, которые, вероятнее всего, появились у общего предка сцеплянок и высших растений (не раньше и не позже). Оказалось, что эти новые гены можно грубо разделить на четыре группы. Во-первых, это факторы транскрипции, то есть регуляторы активности других генов. Во-вторых, это гены, продукты которых вовлечены в гормональную сигнализацию; тут надо учитывать, что гормоны растений — фитогормоны — ничуть не менее разнообразны, чем гормоны животных, и роль их в регуляции роста и развития огромна. В-третьих, это гены, продукты которых участвуют или в создании клеточной стенки, или в ее перестройке (ремоделировании). Клеточные стенки — самая прочная часть растительного организма, его механическая основа. И четвертая группа — гены, кодирующие антистрессорные белки, которые облегчают реакцию клеток на экстремальные условия: например, на высыхание.
Приобретения оказались достаточно масштабными. Например, у общего предка высших растений и сцеплянок, возможно, появилось сразу три новых семейства факторов транскрипции (GRAS, KNOX2 и BBR/BPC). Это много: сравнительная геномика нас учит, что даже в масштабах сотен миллионов лет новые генные семейства появляются не так уж часто, а когда появляются, это обычно связано с появлением нового плана строения или образа жизни (см. Дупликация гомеобоксных генов могла быть одной из причин кембрийского взрыва, «Элементы», 13.02.2015). Какой же новый образ жизни освоил общий предок сцеплянок и высших растений?
Здесь стоит присмотреться к «главным героям» геномного исследования. Мезотениум и спироглея — самые примитивные современные сцеплянки (в данном случае слово «примитивный» обозначает эволюционную ветвь, рано отошедшую от общего ствола). Спироглея — одноклеточная водоросль, мезотениум — нитчатая, хотя и ее клетки тоже вполне могут жить поодиночке (см. Mesotaenium). Но самое интересное, что обе они живут в основном на суше, хотя и во влажных, конечно, местах: на почве, на камнях, на мху. Не архаичный ли это признак? Авторы обсуждаемой работы уверены, что общий предок сцеплянок и высших растений жил в субаэральных условиях — то есть, попросту говоря, в наземных. Это прекрасно объясняет его новшества, особенно приобретение генов, влияющих на структуру клеточной стенки и на устойчивость к высыханию.
А еще это означает, что современные сцеплянковые водоросли, ведущие полностью водный образ жизни — которых все же большинство — вернулись в воду вторично. Но почему бы и нет?
Еще в 1980 году американские ботаники Ледьярд Стеббинс (George Ledyard Stebbins) и Джерри Хилл (Jerry Hill) опубликовали статью под названием: «Выходили ли многоклеточные растения на сушу?» (G. L. Stebbins, G. J. C. Hill, 1980. Did multicellular plants invade the land?). Для многих этот вопрос был неожиданным. Традиционно было принято думать, что какие-то сложные многоклеточные водоросли, имевшие зачатки листьев и жившие на пресноводном мелководье, сначала приспособились поднимать над водой стебель, а потом постепенно освоили незатопляемую сушу и стали высшими растениями (см., например: В. Л. Комаров, 1961. Происхождение растений). Но данные, накопившиеся к концу 1970-х годов, заставили Стеббинса и Хилла сделать вывод, что этот классический сценарий, собственно, ни из чего не следует.
Внешне высшие растения больше всего напоминают харовые водоросли. Но это сходство — явно конвергентное, то есть приобретенное независимо. Все остальные стрептофиты устроены крайне просто: в этой группе нет никаких намеков на расчлененные многоклеточные тела, которые могли бы сначала «высунуть голову» из воды, а потом распространиться на сухой берег. Гораздо вероятнее, что стрептофиты начинали осваивать сушу, будучи еще одноклеточными. Поэтому Стеббинс и Хилл считали, что высшие растения (вернее, их предки) стали многоклеточными уже на суше (рис. 3). И мы видим, что современные геномные исследования подтверждают этот вывод. Действительно, все указывает на то, что общие предки сцеплянок и высших растений были одноклеточными и наземными. Только потом некоторые их потомки вернулись в воду, причем успев «по дороге» потерять жгутики. Это признак наземной адаптации: жгутиковые подвижные стадии очень полезны в воде, а вот на суше без них можно и обойтись.
Впрочем, у высших растений потеря жгутиков не произошла (вернее, произошла, но на целую геологическую эру позже). По этому признаку они архаичнее сцеплянок.

Рис. 3. Сценарий эволюции стрептофит, соответствующий представлениям Стеббинса и Хилла. Они считали, что исходно наземными были не только общие предки сцеплянок и высших растений, но и вообще все стрептофиты — с многократными возвратами в воду. Харовые и десмидиевые водоросли — примеры таких вторичных возвратов. Иллюстрация из статьи: J. Harholt et al., 2016. Why plants were terrestrial from the beginning, с изменениями
Длинная рука ГПГ
Особая проблема связана с происхождением одной группы регуляторных генов — упоминавшихся выше факторов транскрипции GRAS.
Это большое семейство, у высших растений в него входят десятки генов. Любой регуляторный ген — это переключатель, который потенциально может регулировать что угодно. Реально, конечно, у большинства групп регуляторных генов есть какие-то свои типичные функции, но в ходе эволюции они могут довольно легко расширяться и меняться. GRAS-гены участвуют в росте, развитии, адаптации к стрессовым факторам, а также во взаимодействии растений со своими симбионтами (например, с микоризными грибами и с клубеньковыми бактериями). У сцеплянковых водорослей GRAS-гены тоже есть — в частности, у мезотениума и спироглеи это генное семейство представлено прекрасно. Проблема в том, что поиск по генетическим базам данных не обнаружил гомологов GRAS-генов ни у каких других эукариот. Зато обнаружил у бактерий. Все известные на данный момент обладатели GRAS-генов — это высшие растения, сцеплянковые водоросли и пять групп бактерий: дельта-протеобактерии, бактероиды, цианобактерии, актиномицеты и грамположительные бактерии. Поразительно, насколько эти группы разные! Такое широкое распространение GRAS-генов среди совершенно различных бактерий наверняка означает, что сами бактерии активно обменивались этими генами путем горизонтального переноса (ГПГ). Как известно, у бактерий горизонтальный перенос генов — дело обычное (см., например: В эволюции бактерий горизонтальный генетический обмен играет ту же роль, что и половое размножение у высших организмов, «Элементы», 09.04.2012). Очевидно, GRAS-гены были им полезны для регуляции работы генома, в частности, через метилирование ДНК (см. GRAS domain family).
Но откуда взял GRAS-гены общий предок высших растений и сцеплянок? Унаследовать их от более древних эукариот он не мог, потому что (насколько сейчас известно) ни у каких других эукариот этих генов просто нет. Скорее всего, они были получены в результате горизонтального переноса от каких-то почвенных бактерий (см. Гены бактерий помогли водорослям выбраться на сушу, «Элементы», 06.12.2019). От каких именно — сказать трудно, потому что «внутрибактериальный» горизонтальный перенос очень сильно запутал картину. Но это и не так уж важно. Еще Стеббинс и Хилл писали, что некоторые особенности стрептофитов могут объясняться адаптацией к жизни на влажной почве. Вот они и адаптировались, в том числе и таким способом.
Горизонтальный перенос генов от бактерий к эукариотам — явление далеко не такое частое, как перенос между разными бактериями, но и не слишком редкое (см., например: Получение генов пектиназ от протеобактерий резко ускорило видообразование палочников, «Элементы», 15.06.2016). У сцеплянок и высших растений это, похоже, не единственное генное семейство, приобретенное таким способом. Для скептического отношения тут нет оснований, не стоит только впадать в другую крайность и рассматривать горизонтальный перенос генов как универсальный движитель эволюции. Если общий предок сцеплянок и высших растений, как это предполагается, получил новые семейства генов от почвенных бактерий, то он сначала вышел на сушу и только потом нахватался генов от микроорганизмов, с которыми там пришлось иметь дело. То есть первопричиной освоения растениями суши эти гены не были.
От растений к животным
Недавно два известных биолога, палеонтолог Филип Донохью (Philip C. J. Donoghue) и специалист по молекулярной биологии развития Йорди Папс (Jordi Paps Montserrat), выпустили статью, предлагающую своего рода синтетический взгляд на современное состояние проблемы происхождения высших растений. Стоит отметить, что вышла она в журнале Current Biology, который вообще часто публикует полезные обзорные статьи по актуальным проблемам, краткие, но очень содержательные. Донохью и Папс рассматривают данные о происхождении адаптаций, определивших облик высших растений — и предлагают свои выводы.
О том же самом свидетельствует прослеженная биоинформатикой история нескольких других групп генов, влияющих на структуру клеточной стенки, на гормональную сигнализацию и на устойчивость к стрессовым факторам — таким, как высыхание и чрезмерная освещенность (для наземных водорослей последнее немаловажно). Даже у одного из самых примитивных стрептофит — клебсормидиевой водоросли Klebsormidium flaccidum — при ближайшем рассмотрении обнаружился набор сигнальных и антистрессовых систем, которые раньше считались уникальными для высших растений (K. Hori et al., 2014. Klebsormidium flaccidum genome reveals primary factors for plant terrestrial adaptation). Клебсормидиум — морфологически очень простая водоросль, ее колонии имеют форму линейных нитей, и нет никаких указаний на то, что ее предки когда-нибудь были устроены более сложно. Живет она не только в воде, но и в наземных местообитаниях — на коре, на почве, на камнях. Так что, вероятно, Стеббинс и Хилл были в какой-то мере правы: у стрептофит с самого начала была склонность к наземной жизни. У хлорофит такое тоже встречается (см. Trentepohlia), только реже. Возможно, что в эволюции стрептофит переходы из воды на сушу и обратно происходили много раз.
В современной эволюционной биологии вошло в оборот понятие «инструментарий развития» (developmental toolkit). Имеется в виду набор генов и белков (в первую очередь регуляторных), образующих элементную базу для сборки более-менее сложного организма. Инструментарий развития всегда многофункционален, поэтому архитектура организмов, построенных на основе близких наборов генов, может достаточно сильно отличаться. Вероятно, можно сказать, что на регуляторные гены, сигнальные пути и другие элементы инструментария развития распространяется сформулированный в свое время физиологами и морфологами принцип мультифункциональности, дополнительным по отношению к которому является принцип множественного обеспечения функций (см. С. П. Маслов, 1980. Ограничение возможностей гомеостаза мультифункциональностью и главные пути его обхода). Возможности варьирования функций в мультифункциональной системе очень широки. Молекулярный «инструмент», необходимый для развития многоклеточного организма, вполне может пригодиться одноклеточному — и наоборот.
Инструментарий развития, обеспечивающий формирование сложного организма высших растений, формировался в ходе эволюции постепенно, во много этапов, и некоторая его часть возникла уже у одноклеточных водорослей. Сравнительная геномика демонстрирует это вполне наглядно. И это — не особенность, свойственная лишь эволюции растений. Донохью и Папс отмечают, что сценарии происхождения высших растений и многоклеточных животных выглядят с этой точки зрения очень похоже.
Образно говоря, Holozoa шли к многоклеточности широким фронтом. Сравнительная геномика демонстрирует, что при достижении многоклеточности у них произошло не столько «изобретение» совершенно новых генов, сколько кооптирование древних генов для новых задач (см. A. Sebe-Pedros et al., 2017. The origin of Metazoa: a unicellular perspective). Довольно многие гены, считавшиеся раньше уникальными для многоклеточных животных, в действительности оказались свойственными всем Holozoa, а иногда и всем Opisthokonta. В генетический репертуар одноклеточного предка животных совершенно точно уже входили многие гены, функции которых у животных связаны с клеточной адгезией, межклеточной сигнализацией и регуляцией транскрипции (см., например: У одноклеточных организмов есть ген, способный управлять развитием хорды, «Элементы», 25.10.2013). Но в разных эволюционных ветвях эти гены реализовали свои возможные функции по-разному. В некоторых ветвях некоторые гены вообще исчезли, подобно тому, как у сцеплянковых водорослей исчезли гены, кодирующие белки жгутиков и часть факторов иммунитета. В истории происхождения многоклеточных животных и высших растений явно просматривается общий сюжет. Общий, как выражаются иные мыслители, паттерн.
Пульсирующая морфологическая вселенная
Рассмотрим эволюционную ветвь стрептофит. Пройдя адаптивную радиацию, она разделилась на шесть-семь ветвей следующего порядка (они были перечислены выше). Представители этих ветвей, с одной стороны, пополняют свой инструментарий развития, а с другой — морфологически усложняются. Связь между этими событиями, конечно, есть, но благодаря принципу мультифункциональности она очень непрямая: Донохью и Папс так и говорят, что между геномной и морфологической эволюцией наблюдается разрыв (disconnect). Это — фон, на котором происходит становление высших растений.
Высшие растения иначе называются эмбриофитами, потому что у них есть многоклеточный зародыш. Этот и некоторые другие признаки маркируют выход на новый уровень организации (что бы это словосочетание ни обозначало). Если воспользоваться классической терминологией, перед нами типичный ароморфоз.
Но вот что интересно. Разные ветви стрептофит эволюционировали в сторону нового уровня организации в большой мере параллельно. Самые продвинутые — харовые водоросли — успели стать во многом похожими на высшие растения: у них есть не только тело, четко дифференцированное на органы, но и аналоги архегониев — женских органов полового размножения, наличие которых всегда считалось одним из ключевых признаков высших растений (многоклеточный зародыш последних находится именно внутри архегония). Но успешный переход на уровень специализированных наземных многоклеточных организмов совершили вовсе не харовые водоросли. И не колеохетовые, у которых тоже есть многоклеточное тело. Его совершила эволюционная линия, начинающаяся с одноклеточного общего предка эмбриофит и сцеплянок. К переходу преадаптировались многие, но удался он лишь одной группе, причем не самой морфологически сложной, а совсем наоборот — самой простой.
Получается, что в морфологических координатах ароморфоз отнюдь не выглядит как постепенный, инерционный, закономерный процесс. Пусть преадаптация к нему и идет «широким фронтом» (просто за счет обитания многих групп в промежуточной зоне), но главный прорыв удается совершить единицам. «Много званых, но мало избранных».
Более того, похоже, что в нашем случае прорыв в новую среду произошел через упрощение структуры. Ясно, что сцеплянковые водоросли сейчас упрощены по сравнению со своими предками (теми, кто еще не относился к этой группе). Но и заочно знакомый нам общий предок сцеплянок и эмбриофит, по мнению Донохью и Папса, вполне мог быть вторично упрощен по сравнению с древними группами, имевшими более сложную морфологию. Исходя из логики эволюционной теории это не так уж удивительно: сбрасывание груза специализаций вполне может расширять адаптивные возможности (см., например: В. А. Красилов, 1986. Нерешенные проблемы теории эволюции). Получается, что при ароморфозе эволюционирующая группа проходит сквозь этакое морфологическое «бутылочное горлышко»: в новую среду она берет с собой только самое необходимое. А вот ветви с пышной морфологией имеют шансы оказаться тупиковыми. Впрочем, мы пока не знаем, насколько эти закономерности общие.
У проблемы происхождения высших растений есть и еще один интересный аспект. Как известно, эукариотные клетки бывают гаплоидными (с одним набором хромосом) или диплоидными (с двумя наборами). Целый организм тоже может быть гаплоидным, а может быть диплоидным. У всех стрептофитовых водорослей (кроме высших растений) организм гаплоидный. Единственная диплоидная стадия в их жизненном цикле — это зигота, которая сразу делится мейозом и распадается на гаплоидные клетки. И одноклеточные десмидиевые водоросли, и нитчатая спирогира, и пышно ветвящаяся хара — все это гаплоидные существа, гаплобионты.
Высшие растения — единственные из стрептофит, у кого возникла многоклеточная диплоидная стадия. Она называется спорофитом, а более архаичная многоклеточная гаплоидная стадия — гаметофитом (см. Современные высшие растения возникли в результате сдвига экспрессии генов, «Элементы», 10.04.2018). У современных высших растений спорофиты резко отличаются от гаметофитов, и у всех, кроме мхов, они в жизненном цикле доминируют. Сложный дифференцированный многоклеточный спорофит — пожалуй, важнейшее приобретение высших растений (рис. 1, фото 6). Стеббинс и Хилл предполагали, что в суровых наземных условиях у спорофита была выше жизнестойкость, аналогично тому, как и сейчас она часто повышается у полиплоидных разновидностей растений (больше чем с двумя хромосомными наборами).
В любом случае очевидно, что многоклеточный спорофит развился из одноклеточного спорофита, то есть из зиготы. И вот это уж точно истинное «бутылочное горлышко»: ведь собственной многоклеточной морфологии у зиготы нет. В этом смысле весь огромный морфологический мир спорофитов возник de novo — подобно тому, как могла бы возникнуть новая физическая вселенная после прохождения через сингулярность (см. также: Расширение белковой вселенной продолжается, «Элементы», 24.05.2010). Разумеется, некоторая преемственность при этом все равно имела место (иначе процесс, о котором мы говорим, просто не был бы эволюцией), но на морфологическом уровне она была сведена к минимуму. Так что крупные эволюционные изменения далеко не всегда полностью непрерывны.