Ассоциативные области




![]()
Все сенсорные проекционные зоны и моторная область коры занимают менее 20% поверхности коры большого мозга. Остальная кора составляет ассоциативную область. Каждая ассоциативная область коры связана мощными связями с несколькими проекционными областями. Считают, что в ассоциативных областях происходит ассоциация разносенсорной информации. В результате формируются сложные элементы сознания.
Ассоциативные области мозга у человека наиболее выражены в лобной, теменной и височной долях.
Каждая проекционная область коры окружена ассоциативными областями. Нейроны этих областей чаще полисенсорны, обладают большими способностями к обучению. Так, в ассоциативном зрительном поле 18 – число нейронов, «обучающихся» условнорефлекторной реакции на сигнал, составляет более 60% от числа фоновоактивных нейронов. Для сравнения: таких нейронов в проекционном поле 17, всего 10 – 12%.
Повреждение поля 18 приводит к зрительной агнозии. Больной видит, обходит предметы, но не может их назвать.
Полисенсорность нейронов ассоциативной области коры обеспечивает их участие в интеграции сенсорной информации, взаимодействие сенсорных и моторных областей коры.
В теменной ассоциативной области коры формируются субъективные представления об окружающем пространстве, о нашем теле. Это становится возможным благодаря сопоставлению соматосенсорной, проприоцептивной и зрительной информации.
Лобные ассоциативные поля имеют связи с лимбическим отделом мозга и участвуют в организации программ действия при реализации сложных двигательных поведенческих актов.
Первой и наиболее характерной чертой ассоциативных областей коры является мультисенсорность их нейронов, причем сюда поступает не первичная, а достаточно обработанная информация с выделением биологической значимости сигнала. Это позволяет формировать программу целенаправленного поведенческого акта.
Вторая особенность ассоциативной области коры заключается в способности к пластическим перестройкам в зависимости от значимости поступающей сенсорной информации.
Третья особенность ассоциативной области коры проявляется в длительном хранении следов сенсорных воздействий. Разрушение ассоциативной области коры приводит к грубым нарушениям обучения, памяти.
Распределение функций по областям мозга не является абсолютным. Установлено, что практически все области мозга имеют полисенсорные нейроны, т. е. нейроны, реагирующие на различные раздражения. Например, при повреждении поля 17 зрительной области его функцию могут выполнять поля 18 и 19. Кроме того, разные двигательные эффекты раздражения одного и того же двигательного пункта коры наблюдаются в зависимости от текущей моторной деятельности.
Если операцию удаления одной из зон коры провести в раннем детском возрасте, когда распределение функций еще не жестко закреплено, функция утраченной области практически полностью восстанавливается, т. е. в коре имеются проявления механизмов динамической локализации функций, позволяющих компенсировать функционально и анатомически нарушенные структуры.
Важной особенностью коры большого мозга является ее способность длительно сохранять следы возбуждения.
Следовые процессы в спинном мозге после его раздражения сохраняются в течение секунды; в подкорково-стволовых отделах (в форме сложных двигательно-координаторных актов, доминантных установок, эмоциональных состояний) длятся часами; в коре мозга следовые процессы могут сохраняться по принципу обратной связи в течение всей жизни. Это свойство придает коре исключительное значение в механизмах ассоциативной переработки и хранения информации, накопления базы знаний.
Электрические проявления активности коры большого мозга.Оценка функционального состояния коры большого мозга человека является трудной и до настоящего времени нерешенной задачей. Одним из признаков, косвенно свидетельствующем о функциональном состоянии структур головного мозга, является регистрация в них колебаний электрических потенциалов.
Если электрод располагается на поверхности кожи головы, то регистрируется суммарная активность как коры, так и подкорковых структур. Это проявление активности называется электроэнцефалограммой (ЭЭГ).

Все виды активности мозга в динамике подвержены усилению и ослаблению и сопровождаются определенными ритмами электрических колебаний. У человека в покое при отсутствии внешних раздражений преобладают медленные ритмы изменения состояния коры мозга, что на ЭЭГ находит отражение в форме так называемого альфа-ритма, частота колебаний которого составляет 8 – 13 в секунду, а амплитуда – приблизительно 50 мкВ.
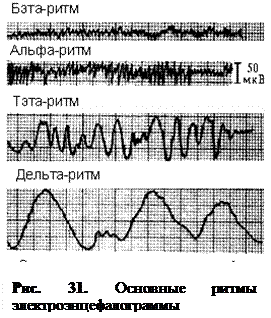 Переход человека к активной деятельности приводит к смене альфа-ритма на более быстрый бета-ритм, имеющий частоту колебаний 14 – 30 в секунду, амплитуда которых составляет 25 мкВ.
Переход человека к активной деятельности приводит к смене альфа-ритма на более быстрый бета-ритм, имеющий частоту колебаний 14 – 30 в секунду, амплитуда которых составляет 25 мкВ.
Переход от состояния покоя к состоянию сосредоточенного внимания или ко сну сопровождается развитием более медленного тета-ритма (4 – 8 колебаний в секунду) или дельта-ритма (0,5 – 3,5 колебаний в секунду). Амплитуда медленных ритмов составляет 100–300 мкВ (рис. 31). В норме у бодрствующего человека дельта-ритм не проявляется.
В течение первого года жизни формируется строго ритмическая электрическая активность частотой 5 Гц с фокусом в затылочной области коры, которая может рассматриваться как аналог альфа-ритма взрослого человека. С возрастом отмечается прогрессивное учащение альфа-ритма, появление и стабилизация его в центральных областях коры. Формирование основного ритма электрической активности, отражающее морфофункциональное созревание нейронного аппарата коры больших полушарий, заканчивается к 16 – 18-летнему возрасту. Параллельно с изменениями ЭЭГ, обусловленными функциональным созреванием коры больших полушарий, отмечается уменьшение выраженности подкорковых знаков в ЭЭГ (тета-волн, билатеральных пароксизмальных разрядов, усиленных неспецифических ответов). Ослабление подкорковых знаков в ЭЭГ с возрастом можно объяснить усилением тормозных влияний созревающей коры на подкорковые структуры.
Таким образом, выявляются определенные этапы функционального созревания коры и подкорковых структур мозга, специфика их взаимных влияний в различные возрастные периоды.
Межполушарная ассиметрия. Результаты исследований различных авторов указывают на то, что между правым и левым полушарием головного мозга существуют анатомические различия (Хомская Е. Д., 2005): размеры, глубина и протяженность извилин и борозд у правшей слева больше, чем у левшей; различается длина полушарий (например, более чем в 50% случаев левое полушарие длиннее правого); имеются различия в размерах нейронов (например, нейроны 3-го и 4-го слоев в 44 и 45 полях левого полушария больше, чем в правом).
Результаты физиологических исследований показали, что существуют различия в ЭЭГ правого и левого полушарий. Многие авторы считают, что при интеллектуальной деятельности межполушарная асимметрия по показателям альфа-ритма усиливается. Также было отмечено, что при выполнении вербальных и невербальных заданий задействуются различные паттерны межполушарной асимметрии (по показателям альфа- и бета-ритмов): при переходе от невербальных заданий к вербальным происходит смена правополушарного доминирования на левополушарное.
Проведенное американскими учеными исследование показало, что дети с более симметричными долями головного мозга хуже справляются с тестами на способности к языкам, т. е. хуже читают и разговаривают.
Благодаря нейропсихологическим исследованиям было установлено, что полушария головного мозга различаются по функциональной специализации. В настоящее время считается доказанным, что с функциями левого и правого полушария у человека связаны два типа мышления – абстрактно-логическое и пространственно-образное. Эти типы мышления имеют ряд синонимов. По В. Ротенбергу:
— вербальное и невербальное (поскольку абстрактно-логическое мышление в отличие от образного базируется на способности к продуцированию речи);
— аналитическое и синтетическое (поскольку с помощью логического мышления осуществляется анализ предметов и явлений, тогда как образное мышление обеспечивает цельность восприятия);
— дискретное и симультанное (поскольку с помощью логического мышления осуществляется ряд последовательных операций, тогда как образное мышление обладает способностью к одномоментному восприятию и оценке объекта).
Также было показано, что правополушарное мышление, создающее специфический пространственно-образный контекст, имеет решающее значение для творчества. Так, при органическом поражении левого полушария мозга у художников и музыкантов практически не страдают их артистические способности, а иногда даже повышается уровень эстетической выразительности творчества. При этом поражения правого полушария способны привести к полной утрате способности к творчеству. Вместе с этим все еще не выясненными остаются вопросы соотношения ведущей руки и ведущего речевого полушария, связи межполушарной асимметрии с эмоциональной сферой и такими психическими познавательными процессами, как память и воображение.
Следует отметить, что, несмотря на богатый фактический материал и активно ведущиеся исследования, единой теории, объясняющей все аспекты межполушарной функциональной асимметрии, до сих пор не существует.
Функции ассоциативных областей коры полушарий большого мозга
![]()
![]()
Все сенсорные проекционные зоны и моторная область коры занимают менее 20% поверхности коры большого мозга (см. рис. 4.14). Остальная кора составляет ассоциативную область. Каждая ассоциативная область коры связана мощными связями с несколькими проекционными областями. Считают, что в ассоциативных областях происходит ассоциация разносенсорной информации. В результате формируются сложные элементы сознания.
Ассоциативные области мозга у человека наиболее выражены в лобной, теменной и височной долях.
Каждая проекционная область коры окружена ассоциативными областями. Нейроны этих областей чаще полисенсорны, обладают большими способностями к обучению. Так, в ассоциативном зрительном поле 18 число нейронов, «обучающихся» условнорефлекторной реакции на сигнал, составляет более 60% от числа фоновоактивных нейронов. Для сравнения: таких нейронов в проекционном поле 17 всего 10—12%.
Повреждение поля 18 приводит к зрительной агнозии. Больной видит, обходит предметы, но не может их назвать.
Полисенсорность нейронов ассоциативной области коры обеспечивает их участие в интеграции сенсорной информации, взаимодействие сенсорных и моторных областей коры.
В теменной ассоциативной области коры формируются субъективные представления об окружающем пространстве, о нашем теле. Это становится возможным благодаря сопоставлению соматосенсорной, проприоцептивной и зрительной информации.
Лобные ассоциативные поля имеют связи с лимбическим отделом мозга и участвуют в организации программ действия при реализации сложных двигательных поведенческих актов.
Первой и наиболее характерной чертой ассоциативных областей коры является мультисенсорность их нейронов, причем сюда поступает не первичная, а достаточно обработанная информация с выделением биологической значимости сигнала. Это позволяет формировать программу целенаправленного поведенческого акта.
Вторая особенность ассоциативной области коры заключается в способности к пластическим перестройкам в зависимости от значимости поступающей сенсорной информации.
Третья особенность ассоциативной области коры проявляется в длительном хранении следов сенсорных воздействий. Разрушение ассоциативной области коры приводит к грубым нарушениям обучения, памяти. Речевая функция связана как с сенсорной, так и с двигательной системами. Корковый двигательный центр речи расположен в заднем отделе третьей лобной извилины (поле 44) чаще левого полушария и был описан вначале Даксом (1835), а затем Брока (1861).
Слуховой центр речи расположен в первой височной извилине левого полушария (поле 22). Этот центр был описан Вернике (1874). Моторный и слуховой центры речи связаны между собой мощным пучком аксонов.
Речевые функции, связанные с письменной речью, — чтение, письмо — регулируются ангулярной извилиной зрительной области коры левого полушария мозга (поле 39).
При поражении моторного центра речи развивается моторная афазия; в этом случае больной понимает речь, но сам говорить не может. При поражении слухового центра речи больной может говорить, излагать устно свои мысли, но не понимает чужой речи, слух сохранен, но больной не узнает слов. Такое состояние называется сенсорной слуховой афазией. Больной часто много говорит (логорея), но речь его неправильная (аграмматизм), наблюдается замена слогов, слов (парафазии).
Поражение зрительного центра речи приводит к невозможности чтения, письма.
Изолированное нарушение письма — аграфия, возникает также в случае расстройства функции задних отделов второй лобной извилины левого полушария.
В височной области расположено поле 37, которое отвечает за запоминание слов. Больные с поражениями этого поля не помнят названия предметов. Они напоминают забывчивых людей, которым необходимо подсказывать нужные слова. Больной, забыв название предмета, помнит его назначения, свойства, поэтому долго описывает их качества, рассказывает, что делают этим предметом, но назвать его не может. Например, вместо слова «галстук» больной, глядя на галстук, говорит: «это то, что надевают на шею и завязывают специальным узлом, чтобы было красиво, когда идут в гости».
Распределение функций по областям мозга не является абсолютным. Установлено, что практически все области мозга имеют полисенсорные нейроны, т. е. нейроны, реагирующие на различные раздражения. Например, при повреждении поля 17 зрительной области его функцию могут выполнять поля 18 и 19. Кроме того, разные двигательные эффекты раздражения одного и того же двигательного пункта коры наблюдаются в зависимости от текущей моторной деятельности.
Важной особенностью коры большого мозга является ее способность длительно сохранять следы возбуждения.
Следовые процессы в спинном мозге после его раздражения сохраняются в течение секунды; в подкорково-стволовых отделах (в форме сложных двигательно-координаторных актов, доминантных установок, эмоциональных состояний) длятся часами; в коре мозга следовые процессы могут сохраняться по принципу обратной связи в течение всей жизни. Это свойство придает коре исключительное значение в механизмах ассоциативной переработки и хранения информации, накопления базы знаний.
Сохранение следов возбуждения в коре проявляется в колебаниях уровня ее возбудимости; эти циклы длятся в двигательной области коры 3—5 мин, в зрительной — 5—8 мин.
Основные процессы, происходящие в коре, реализуются двумя состояниями: возбуждением и торможением. Эти состояния всегда реципрокны. Они возникают, например, в пределах двигательного анализатора, что всегда наблюдается при движениях; они могут возникать и между разными анализаторами. Тормозное влияние одного анализатора на другие обеспечивает сосредоточенность внимания на одном процессе.
Реципрокные отношения активности очень часто наблюдаются в активности соседних нейронов.
Отношение между возбуждением и торможением в коре проявляется в форме так называемого латерального торможения. При латеральном торможении вокруг зоны возбуждения формируется зона заторможенных нейронов (одновременная индукция) и она по протяженности, как правило, в два раза больше зоны возбуждения. Латеральное торможение обеспечивает контрастность восприятия, что в свою очередь позволяет идентифицировать воспринимаемый объект.
Помимо латерального пространственного торможения, в нейронах коры после возбуждения всегда возникает торможение активности и наоборот, после торможения — возбуждение — так называемая последовательная индукция.
В тех случаях когда торможение не в состоянии сдерживать возбудительный процесс в определенной зоне, возникает иррадиация возбуждения по коре. Иррадиация может происходить от нейрона к нейрону, по системам ассоциативных волокон I слоя, при этом она имеет очень малую скорость — 0,5—2,0 м/с. В другом случае иррадиация возбуждения возможна за счет аксонных связей пирамидных клеток III слоя коры между соседними структурами, в том числе между разными анализаторами. Иррадиация возбуждения обеспечивает взаимоотношение состояний систем коры при организации условнорефлекторного и других форм поведения.
Моторные области
Впервые Фритч и Гитциг (1870) показали, что раздражение передней центральной извилины мозга (поле 4) вызывает двигательную реакцию. В то же время признано, что двигательная область является анализаторной.
Спереди от передней центральной извилины лежат премоторные поля 6 и 8. Они организуют не изолированные, а комплексные, координированные, стереотипные движения. Эти поля также обеспечивают регуляцию тонуса гладкой мускулатуры, пластический тонус мышц через подкорковые структуры.
В реализации моторных функций принимают участие также вторая лобная извилина, затылочная, верхнетеменная области.
Двигательная область коры, как никакая другая, имеет большое количество связей с другими анализаторами, чем, видимо, и обусловлено наличие в ней значительного числа полисенсорных нейронов.
Ассоциативные области
Все сенсорные проекционные зоны и моторная область коры занимают менее 20% поверхности коры большого мозга (см. рис. 4.14). Остальная кора составляет ассоциативную область. Каждая ассоциативная область коры связана мощными связями с несколькими проекционными областями. Считают, что в ассоциативных областях происходит ассоциация разносенсорной информации. В результате формируются сложные элементы сознания.
Ассоциативные области мозга у человека наиболее выражены в лобной, теменной и височной долях.
Каждая проекционная область коры окружена ассоциативными областями. Нейроны этих областей чаще полисенсорны, обладают большими способностями к обучению. Так, в ассоциативном зрительном поле 18 число нейронов, «обучающихся» условнорефлекторной реакции на сигнал, составляет более 60% от числа фоновоактивных нейронов. Для сравнения: таких нейронов в проекционном поле 17 всего 10—12%.
Повреждение поля 18 приводит к зрительной агнозии. Больной видит, обходит предметы, но не может их назвать.
Полисенсорность нейронов ассоциативной области коры обеспечивает их участие в интеграции сенсорной информации, взаимодействие сенсорных и моторных областей коры.
В теменной ассоциативной области коры формируются субъективные представления об окружающем пространстве, о нашем теле. Это становится возможным благодаря сопоставлению соматосенсорной, проприоцептивной и зрительной информации.
Лобные ассоциативные поля имеют связи с лимбическим отделом мозга и участвуют в организации программ действия при реализации сложных двигательных поведенческих актов.
Первой и наиболее характерной чертой ассоциативных областей коры является мультисенсорность их нейронов, причем сюда поступает не первичная, а достаточно обработанная информация с выделением биологической значимости сигнала. Это позволяет формировать программу целенаправленного поведенческого акта.
Вторая особенность ассоциативной области коры заключается в способности к пластическим перестройкам в зависимости от значимости поступающей сенсорной информации.
Третья особенность ассоциативной области коры проявляется в длительном хранении следов сенсорных воздействий. Разрушение ассоциативной области коры приводит к грубым нарушениям обучения, памяти. Речевая функция связана как с сенсорной, так и с двигательной системами. Корковый двигательный центр речи расположен в заднем отделе третьей лобной извилины (поле 44) чаще левого полушария и был описан вначале Даксом (1835), а затем Брока (1861).
Слуховой центр речи расположен в первой височной извилине левого полушария (поле 22). Этот центр был описан Вернике (1874). Моторный и слуховой центры речи связаны между собой мощным пучком аксонов.
При поражении моторного центра речи развивается моторная афазия; в этом случае больной понимает речь, но сам говорить не может. При поражении слухового центра речи больной может говорить, излагать устно свои мысли, но не понимает чужой речи, слух сохранен, но больной не узнает слов. Такое состояние называется сенсорной слуховой афазией. Больной часто много говорит (логорея), но речь его неправильная (аграмматизм), наблюдается замена слогов, слов (парафазии).
Поражение зрительного центра речи приводит к невозможности чтения, письма.
В височной области расположено поле 37, которое отвечает за запоминание слов. Больные с поражениями этого поля не помнят названия предметов. Они напоминают забывчивых людей, которым необходимо подсказывать нужные слова. Больной, забыв название предмета, помнит его назначения, свойства, поэтому долго описывает их качества, рассказывает, что делают этим предметом, но назвать его не может. Например, вместо слова «галстук» больной, глядя на галстук, говорит: «это то, что надевают на шею и завязывают специальным узлом, чтобы было красиво, когда идут в гости».
Распределение функций по областям мозга не является абсолютным. Установлено, что практически все области мозга имеют полисенсорные нейроны, т. е. нейроны, реагирующие на различные раздражения. Например, при повреждении поля 17 зрительной области его функцию могут выполнять поля 18 и 19. Кроме того, разные двигательные эффекты раздражения одного и того же двигательного пункта коры наблюдаются в зависимости от текущей моторной деятельности.
Если операцию удаления одной из зон коры провести в раннем детском возрасте, когда распределение функций еще не жестко закреплено, функция утраченной области практически полностью восстанавливается, т. е. в коре имеются проявления механизмов динамической локализации функций, позволяющих компенсировать функционально и анатомически нарушенные структуры.
Важной особенностью коры большого мозга является ее способность длительно сохранять следы возбуждения.
Следовые процессы в спинном мозге после его раздражения сохраняются в течение секунды; в подкорково-стволовых отделах (в форме сложных двигательно-координаторных актов, доминантных установок, эмоциональных состояний) длятся часами; в коре мозга следовые процессы могут сохраняться по принципу обратной связи в течение всей жизни. Это свойство придает коре исключительное значение в механизмах ассоциативной переработки и хранения информации, накопления базы знаний.
Основные процессы, происходящие в коре, реализуются двумя состояниями: возбуждением и торможением. Эти состояния всегда реципрокны. Они возникают, например, в пределах двигательного анализатора, что всегда наблюдается при движениях; они могут возникать и между разными анализаторами. Тормозное влияние одного анализатора на другие обеспечивает сосредоточенность внимания на одном процессе.
Реципрокные отношения активности очень часто наблюдаются в активности соседних нейронов.
Отношение между возбуждением и торможением в коре проявляется в форме так называемого латерального торможения. При латеральном торможении вокруг зоны возбуждения формируется зона заторможенных нейронов (одновременная индукция) и она по протяженности, как правило, в два раза больше зоны возбуждения. Латеральное торможение обеспечивает контрастность восприятия, что в свою очередь позволяет идентифицировать воспринимаемый объект.
Наряду с иррадиацией возбуждения, которое происходит за счет импульсной передачи активности, существует иррадиация состояния торможения по коре. Механизм иррадиации торможения заключается в переводе нейронов в тормозное состояние под влиянием импульсов, приходящих из возбужденных участков коры, например, из симметричных областей полушарий.
Способность длительно сохранять следы возбуждения а следовые процессы в течение всей жизни
Базальные ганглии — это структуры переднего мозга, хвостатое ядро, скорлупа чечевицеобразного тела или просто скорлупа, бледный шар, субталамическое ядро, функционально к ним относят также черную субстанцию среднего мозга. Считается, что базальные ганглии служат в качестве системы, уточняющей программу выполнения сложных движений.
Кора головного мозга является высшим отделом ЦНС, обеспечивающим на основе врожденных и приобретенных в онтогенезе функций наиболее совершенную организацию поведения организма.
Кора головного мозга имеет ряд морфофункциональных особенностей:
а) многослойность расположения нейронов и модульный принцип организации;
б) соматотопическая локализация рецепторных систем и экранность (распределение внешней рецепции на плоскости нейронального поля коркового конца анализатора);
в) зависимость уровня активности от влияния подкорковых структур и ретикулярной формации и наличие представительства всех функций нижележащих структур ЦНС;
г) цитоархитектоническое распределение на поля и наличие в специфических проекционных сенсорных и моторных системах коры вторичных и третичных полей с превалированием ассоциативных областей коры (наличие специализированных ассоциативных областей коры);
д) динамическая локализация функций, выражающаяся в возможности компенсаций функций утраченных структур коры (перекрытие в коре зон соседних периферических рецептивных полей, способность к иррадиации);
е) возможность длительного сохранения следов раздражения и реципрокная функциональная взаимосвязь возбудимых и тормозных состояний коры (наличие специфической электрической активности).
Значительное место в клеточном составе коры занимают пирамидные нейроны. Пирамидные нейроны имеют разную величину, их дендриты несут большое количество шипиков; аксон, как правило, идет через белое вещество в другие зоны коры или в другие структуры ЦНС. Звездчатые клетки имеют короткие дендриты и короткий аксон, обеспечивающий связи нейронов самой коры. Веретенообразные нейроны обеспечивают вертикальные или горизонтальные взаимосвязи нейронов.
Особенностью коры головного мозга является ее способность длительно сохранять следы возбуждения. В спинном мозге после раздражения следовые процессы сохраняются секунды; в подкорково-стволовых отделах — в форме сложных двигательно-координаторных актов, доминантных установок, эмоциональных состояний эти процессы длятся часами; в коре мозга следовые процессы могут сохраняться в течение всей жизни. Это свойство придает коре исключительное значение в механизмах переработки и хранения информации, накопления базы знаний.
Основные процессы, происходящие в ЦНС, а следовательно и в коре, реализуются двумя состояниями: возбуждения и торможения. Эти состояния всегда реципрокны. Они возникают, например, в пределах двигательного анализатора, что наблюдается всегда при движениях, они могут возникать и между разными анализаторами. Тормозное влияние одного анализатора на другие обеспечивает сужение, сосредоточенность внимания на одном процессе. Реципрокные отношения активности часто наблюдаются в соседних нейронах. Отношение между возбуждением и торможением в коре проявляется в форме так называемого латерального торможения. При латеральном торможении вокруг зоны возбуждения формируется зона заторможенных нейронов, и она, как правило, по протяженности в два раза больше зоны возбуждения. Латеральное торможение обеспечивает контрастность восприятия, что, в свою очередь, позволяет идентифицировать воспринимаемый объект. Помимо латерального пространственного торможения, в коре после возбуждения всегда возникает торможение активности, и наоборот, после торможения — возбуждение (последовательная индукция). В тех случаях, когда торможение не в состоянии сдерживать возбудительный процесс в определенной зоне, возникает иррадиация возбуждения в коре. Иррадиация возбуждения в коре обеспечивает взаимоотношение состояний областей коры при организации условнорефлекторного и других форм поведения. Наряду с иррадиацией возбуждения, которое происходит за счет импульсной передачи активности, существует иррадиация торможения по коре. Механизм иррадиации торможения заключается в переводе нейронов в тормозное состояние, за счет торможения приходящих к ним аксонов, их синапсов.
Каждый нейрон имеет заряд мембраны, при активации нейрона этот заряд генерируется в форме импульсных разрядов, при торможении заряд мембраны чаще увеличивается, и возникает ее гиперполяризация. Глия мозга также имеет заряд мембран. Заряд мембраны нейронов и глии, его динамика, процессы, происходящие в синапсах, дендритах, аксонном холмике, в аксоне — все это постоянно изменяющиеся, разнообразные и разнонаправленные по знаку, по интенсивности, по скорости процессы. Их интегральные характеристики зависят от функционального состояния нервной структуры и определяют суммарно ее электрические показатели.