Почему высота тетануса больше чем высота одиночного мышечного сокращения
Тетанус
Тетаническое сокращение
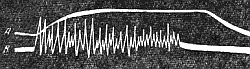
Рис. 142. Одновременная запись тетанического сокращения (А) и токов действия (В) мышцы (по А. Г. Гинецинскому).
Тетанические сокращения мышцы представля ют собой результат суммации одиночных сокращений. Для исследования суммации сокращений наносят на мышцу два одиночных раздражения. Промежуток времени между раздражениями должен быть такой, чтобы второе раздражение подействовало на мышцу раньше, чем она успеет расслабиться после первого сокращения. При этом возможны два случая.
Если второе раздражение поступает, когда мышца уже начала расслабляться после первого сокращения, то на миографической кривой вершина второго сокращения будет отделена от вершины первого небольшим западением кривой ( рис. 143, Б ). Если же второе раздражение действует, когда первое сокращение ещё не дошло до своей вершины, то второе сокращение полностью сливается с первым, образуя вместе с ним единую суммированную вершину ( рис. 143, А ).
Рис. 143. Суммации мышечных сокращений. а — момент первого раздражения; в — момент второго раздражения; 1 — сокращение в ответ на одно раздражение а; 2 — сокращение в ответ на одно раздражение в; 3— суммированное сокращение в ответ на раздражения а и в. А — второе раздражение падает в период сокращения; Б — второе раздражение наносится в период расслабления.
Таким образом, возможны два типа суммации. Рассмотрим в качестве примера, как они происходят в икроножной мышце лягушки. Продолжительность одиночного сокращения этой мышцы в среднем равна при комнатной температуре 0,1 секунды. Если считать, что восходящее и нисх дящее колена сокращения приблизительно равны, то на долю каждого колена приходится 0,05 секунды. Поэтому для воспроизведения на этой мышце первого типа суммации сокращений (неполной суммации) необходимо, чтобы интервал между первым и вторым раздражением был боьше чем 0,05 секунды, а для получения второго типа суммации (так называемой полной суммации) — меньше чем 0,05 секунды.
Гладкий и зубчатый тетанус
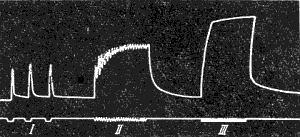
Рис. 144. Сокращения мышцы при разной частоте раздражений. Редкие раздражения вызывают одиночные сокращения (I); более частые — зубчатый тетанус (II), еще более частые — гладкий тетанус (III).
После прекращения тетанического раздражения волокна вначале не полностью расслабляются, и их исходная длина восстанавливается лишь по истечении некоторого времени. Это явление называется послететанической, или остаточной, контрактурой.
Чем быстрее сокращается и расслабляется мышца, тем чаще должны быть раздражения, необходимые для того, чтобы вызвать тетанус. Так, для тетануса белых мышц кролика (быстрых) требуется 30 индукционных ударов в секунду, а для тетануса красных (медленных) мышц того же животного — только 10 ударов в секунду.
Механизм суммации сокращений при тетанусе
Высота тетанического сокращения мышцы значительно превышает максимальную амплитуду ее одиночного сокращения ( рис. 144 ). Объясняется этот факт тем, что при ритмическом раздражении каждая новая волна возбуждения вызывает дополнительное сокращение мышц, которое суммируется с предыдущим. Г. Гельмгольц (1847) назвал этот процесс суперпозицией, т. е. накладыванием сокращении, полагая, что при каждом следующем импульсе мышца укорачивается так, как если бы она была в этот момент расслаблена.
Однако дальнейшие исследования показали, что явления суперпозиции при тетанусе ни в коем случае нельзя сводить к простому складыванию двух механических эффектов (Н. Е. Введенский. 1880). Выяснилось, что эффект двух следующих друг за другом раздражении отнюдь не равен арифметической сумме одиночных сокращений, а, как правило, оказывается то большим, то меньшим данной суммы. Это свидетельствует о том, что способность к новому сокращению после каждой предшествующей вспышки возбуждения значительно изменяется. Н. Е. Введенский объяснял это тем, что каждая вспышка возбуждения и сокращения оставляет след в ткани в виде повышенной возбудимости. Если второе раздражение действует в тот момент, когда вобудимость мышцы повышена, амплитуда второго, а при ритмических раздражениях и амплитуда тетанического сокращения становится больше, чем одиночного.
Изменения состояния мышцы в процессе тетануса можно хорошо наблюдать при непрямом раздражении мышцы. Если непосредственно после окончания тетанического сокращения портняжной мышцы лягушки на нерв нанести одиночное раздражение максимальной силы (т. е. такой силы, которая оказывается достаточной для возбуждения всех нервных волокон), то возникает одиночное сокращение, которое в амплитуде значительно больше, чем оно было до тетануса.
Согласно Е. Б. Бабскому, изменения состояния мышцы, лежащие в основе формирования тетанического сокращения, связаны с освобождением в мышце при одиночном сокращении аденозинтрифосфорной кислоты, которая не успевает полностью расщепиться до начала следующей вспышки возбуждения. Это вещество в очень малых концентрациях оказывает значительное влияние на возбудимость и сократимость мышцы и может способствовать тому, что каждый следующий приходящий к мышце импульс будет давать больший эффект, чем предыдущие.
Почему высота тетануса больше чем высота одиночного мышечного сокращения
1. Роль мышечного сокращения в организации поведенческой деятельности человека. Классификация мышц и их функции. Виды и режимы мышечных сокращений.
2. Гладкие мышцы, их морфологические и физиологические особенности.
3. Одиночное мышечное сокращение, его фазы. Суммация сокращений.
4. Тетаническое сокращение, его виды. Оптимум и пессимум по Введенскому
5. Сократительный аппарат мышечного волокна. Механизм мышечного сокращения.
6. Синапс. Классификация. Особенности строения. Механизм передачи возбуждения в химическом синапсе. Свойства синапсов.
7. Медиатор. Виды медиаторов. Свойства медиаторов.
8. Электрические и тормозные синапсы. Особенности передачи сигнала.
9. Пути фармакологической регуляции синаптической передачи возбуждения.
1.Роль мышечного сокращения в организации поведенческой деятельности человека. Классификация мышц и их функции. Виды и режимы мышечных сокращений.
Общим свойством всего живого и основой активного поведения являетсядвижение.
Органом движения являетсямышечный аппарат, который включает 3 вида мышц: скелетные, гладкие и сердечную мышцы.
Они выполняют следующиефункции:
1. Создание позы и удержание тела в пространстве, преодоление инерции.
1. Двигательная функция внутренних органов (моторная функция кишечника, сократительная функция сердца, обеспечение дыхания за счет сокращения дыхательных мышц).
2. Эффекторный механизм мыслительной (произносимая речь) и поведенческой деятельности.
3. Преобразование химической энергии макроэргических соединений в механическую, тепловую, электрическую энергию.
Составляют 35-40% массы тела, их количество достигает 600.
Состоят из пучков мышечных волокон, заключенных в общую соединительно-тканную оболочку.
Мышечное волокно – это гигантская, многоядерная мышечная клетка (диаметр от 1 до 100 мкм, длина от 5 до 400 мм), содержащая сотнимиофибрилл, которые являются структурной единицей и представляют сократительный аппарат мышечного волокна. Миофибриллы включают актин и миозин.
Скелетные волокна подpазделяются на фазные волокна (они генерируют потенциал действия) и тонические (не способны генерировать распространяющееся возбуждение).
Фазные волокна делятся на быстрые волокна (белые, гликолитические) и медленные волокна (красные, окислительные).
Физические свойства скелетных мышц.
Растяжимость – это способность мышцы изменять свою длину под действием растягивающей силы.
Эластичность – способность мышцы принимать свою первоначальную длину после прекращения действия растягивающей или деформирующей силы.
Сила – определяется максимальным грузом, который мышца в состоянии поднять.
Способность совершать работу – определяется произведением массы поднятого груза на высоту подъема.
Физиологические свойства скелетных мышц: возбудимость, проводимость, сократимость, лабильность.
Для скелетной мышцы характерны три основных режима сокращения:
ИЗОТОНИЧЕСКИЙ – укорочение мышцы без изменения ее тонического напряжения (когда мышце не приходится перемещать груз, например, сокращение мышц языка).
ИЗОМЕТРИЧЕСКИЙ – длина мышечных волокон остается постоянной на фоне увеличения напряжения (попытка поднять непосильный груз)
АУКСОТОНИЧЕСКИЙ – изменениедлины сопровождается изменением напряжения (работа мышцы при выполнении трудовых, спортивных и других двигательных актов).
Для скелетной мышцы характерны двавида сокращений:
ОДИНОЧНОЕ сокращение – возникает при действии одиночным стимулом (раздражителем) непосредственно на мышцу (прямое раздражение), или через иннервирующий ее двигательный нерв (непрямое).
ТЕТАНИЧЕСКОЕ (суммированное) сокращение – длительное сокращение мышцы в ответ на ритмическое раздражение.
(В естественных условиях к скелетной мышце из ЦНС поступают не одиночные импульсы, а серия импульсов, следующих друг за другом с определенными интервалами).
2.Гладкие мышцы, их морфологические и физиологические особенности
Гладкие мышцы находятся:
· во внутренних органах (пишеварительный тракт, мочевой пузырь);
· в сосудах, коже, глазе (мышцы радужной оболочки, цилиарная мышца).
Тонические – не способны развивать «быстрые» сокращения.
Фазно-тонические – способны быстро сокращаться и подразделяются на обладающие автоматией и не обладающие автоматией.
2. Хаотично расположены и окружены соединительной тканью (поэтому лишены поперечной исчерченности).
3. Контактируют друг с другом при помощи нексусов.
4. Сократительный аппарат представлен миофибриллами, состоящими в основном из актина. Миозин представлен только в дисперсной и агрегированной формах.
1. В основе сокращения – процесс превращения энергии АТФ в механическую энергию сокращения.
2. Сокращения медленные с использованием скользящего механизма.
3. Сокращение протекает с малыми энерготратами.
4. Обладают выраженной пластичностью (длительное сохранение измененной длины).
5. Обладают автоматией.
1. Быстрое и сильное растяжение гладких мышц.
2. Химические вещества (особенно гормоны и медиаторы, к которым гладкие мышцы обладают высокой чувствительностью).
Особенности электрических процессов.
1. Потенциал покоя в гладких мышцах меньше, чем в скелетных.
В клетках не обладающих автоматией он стабилен и =– 60–70 мВ.
В клетках, обладающих автоматией, он неустойчивый с колебаниями от –30 до –70 мВ.
Потенциал действия имеет длительный латентный период.
Ниже, чем в скелетных мышцах.
Бывает двух типов: пикоподобная форма и форма «плато».
Связан с повышением проницаемости для ионов Са.
Несколько опережает сокращение.
2. Проведение возбуждения возникает, если приложенный стимул одновременно возбуждает некоторое минимальное количество мышечных клеток.
Может распространяться на соседние мышечные волокна (из-за малого сопротивления в области контактов) распространяется лишь на определенное расстояние, которое зависит от силы раздражителя скорость значительно меньше, чем в скелетной мышце и составляет от 2 до 15 см/с.
3. Одиночное мышечное сокращение, его фазы. Суммация сокращений
Одиночное мышечное сокращение продолжается около 100 мс и развивается по фазам:
Латентный (скрытый) период продолжается до 3 мс и представляет время от начала действия раздражителя до начала видимого ответа (сокращения) мышцы.
Фаза сокращения продолжается 40-50 мс характеризуется укорочением длины мышечного волокна, что связано с увеличением концентрации Сa 2+ в протофибриллярных пространствах и образованием актин-миозиновых связей.
Фаза расслабления продолжается 50-60 мс характеризуется увеличением (восстановлением) длины волокна. Возникает при снижении концентрации Ca 2+ в протофибриллярных пространствах и ослаблениемактин-миозиновых связей.
Если на мышцу наносятся два и более раздражений с интервалом менее продолжительности одиночного сокращения, но более продолжительности рефрактрного периода ПД, то происходит суммация сокращений, в результате которой сократительный эффект усиливается.
Существуетдва типа суммации: частичная и полная
Частичная (или неполная)суммация возникает, если
· интервал между раздражениями меньше продолжительности одиночного мышечного сокращения;
· больше продолжительности фазы сокращения, т.е. если второе раздражение попадает в фазу расслабления.
В результате амплитуда мышечного сокращения возрастает с образованием двух вершин.
Полная суммация возникает, если:
· интервал между раздражениями меньше продолжительности фазы сокращения, но больше продолжительности рефрактерного периода;
· второе раздражение попадает в фазу сокращения.
В результате амплитуда мышечного сокращения изменяется (увеличивается или уменьшается относительно одиночного сокращения) с образованием одной вершины
Увеличение или уменьшение амплитуды связано с изменением возбудимости в процессе возбуждения и зависит от того в какую фазу измененной возбудимости наносится следующее раздражение.
Известно, что в период формирования пикового потенциала возбудимость ткани снижена (фаза абсолютной и фаза относительной рефрактерности). Поэтому, если следующее раздражение будет наноситься в этот период, то амплитуда мышечного сокращения будет снижена.
Период возбуждения в скелетной мышце завершается следовой деполяризацией, продолжающейся от 20 до 40 мс.
В этот период возбудимость, а, следовательно, и сократимость повышена. Поэтому, если следующее раздражение будет приходиться на этот период, то амплитуда мышечного сокращения будет возрастать (тем больше, чем больше повышена возбудимость).
3.Тетаническое сокращение, его виды. Оптимум и пессимум по Введенскому
Различаютдва вида тетануса: зубчатый и гладкий.
В их основе лежат механизмы частичной или полной суммации.
Вид тетанического сокращения определяется Механическим состоянием мышцы в момент повторного возбуждения. Состоянием возбудимости мышцы в момент повторного возбуждения.
Зубчатый тетанус развивается на ряд последовательных раздражений, интервал между которыми больше продолжительности фазы сокращения, но меньше продолжительности одиночного мышечного сокращения (интервал от 100 до 50 мс при частоте раздражений от 10 до 20 Гц).
При этом каждое новое сокращение формируется на фоне не завершившегося расслабления мышцы, образуя новые вершины последующих сокращений («зубцы»). Высота суммарного сокращения зависит от ритма и силы раздражений и определяется исходным уровнем формирования каждого следующего сокращения (чем выше уровень, тем больше амплитуда).
В начале фазы расслабления этот уровень выше, чем в конце.
Гладкий тетанус развивается на ряд последовательных раздражений, интервал между которыми меньше длительности фазы сокращения, но больше продолжительности потенциала действия (интервал от 50 до 5 мс при частоте 20 до 200 Гц).
Каждое новое сокращение формируется на фоне не завершившегося сокращения мышцы, образуя единую, гладкую вершину. Ее высота определяется уровнем измененной возбудимости в процессе возбуждения.
Если каждый следующий раздражитель попадает в фазу экзальтации (повышенной возбудимости), то амплитуда сокращения будет большой.
Если импульсы попадают в период сниженной возбудимости (относительная рефрактерность), то амплитуда будет снижена.
Явление изменения амплитуды в зависимости от возбудимости мышцы объяснил H.Е.Введенский, введя понятие оптимума и пессимума.
Оптимальная частота – максимальная частота раздражений, при которой возникает максимальная амплитуда тетанического ответа.
Пессимум – снижение амплитуды тетанического сокращения при увеличении частоты раздражений (выше оптимальной величины).
Пессимальная частота – максимальная частота (сверх оптимальной), при которой возникает минимальная амплитуда тетанического ответа.
4.Сократительный аппарат мышечного волокна. Механизм мышечного сокращения
Они разделены на чередующиеся участки (диски), которые обладают различными оптическими свойствами.
Диски, обладающие двойным лучепреломлением, получили название анизотропные (А) диски.
Диски, которые не обладают двойным лучепреломлением, названы изотропные (I) диски.
Анизотропные диски в обыкновенном свете выглядят темными и состоят из двух темных полосок, разделенных светлой «H» полоской.
Изотропные диски в обыкновенном свете выглядят светлыми и в середине имеют темную «Z» полоску.
Z полоска – эта тонкая мембрана, которая является продолжением поверхностной мембраны вглубь мышечного волокна.
Она выполняет опорную функцию, поскольку через ее поры проходят протофибриллы.
В зоне Z мембраны также находятся триады или Т-системы триады представляют выпячивания плазматической мембраны с образованием поперечных трубочек в виде ярусов и цисцерн.
Они предсталяют саркоплазматический ретикулум, который содержит высокую концентрацию ионов Ca.
При возбуждении Z мембраны кальций по концентрационному градиенту выходит из саркоплазматического ретикулума в протофибриллярное пространство, вызывая процесс сокращения Активная реабсорбция ионов Са в саркоплазматический ретикулум за счет работы Са-насоса, приводит к расслаблению мышечного волокна.
Протофибриллы включают белковые нитиактина имиозина, а также белки тропонин и тропомиозин.
Нити миозина – это толстые и короткие нити, которые входят только в состав анизотропного диска.
Нити актина – это тонкие и длинные нити, входящие в состав как изотропного, так и анизотропного дисков. Они вставлены между нитями миозина. От них свободна только H-полоска анизотропного диска.
Процесс сокращения происходит в результате скольжения нитей актина относительно нитей миозина, который запускается накоплением Са, при этом образуются актино-миозиновые комплексы (мостики) и нити актина вдвигаются в промежутки между нитями миозина.
Нити актина сближаются друг с другом.
Ширина H-полоски и анизотропных дисков уменьшается, изотропный диск своего не изменяет своего размера.
Механизм мышечного сокращения и расслабления.
2. Возникновение потенциала действие.
3. Проведение возбуждения вдоль клеточной мембраны до Z мембраны, а далее вглубь волокна по трубочкам саркоплазматического ретикулума.
4. Освобождение Са из триад.
5. Диффузия Са к протофибриллам.
6. Взаимодействие Са с тропонином.
7. Конформационное изменение комплекса тропомиозин-тропонин.
8. Освобождение активных центров актина.
9. Присоединение актина к миозину.
10. В присутствии белка актомиозина распад АТФ с освобождением энергии.
11. Скольжение нитей актина относительно миозина.
12. Укорочение миофибриллы.
13. Активация кальциевого насоса.
15. Понижение концентрации свободных ионов Са в саркоплазме.
16. Разрушение актин-миозиновых комплексов.
17. Обратное скольжение нитей актина относительно миозина.
18. Увеличение (восстановление) миофибриллы.
5. Синапс. Классификация. Особенности строения. Механизм передачи возбуждения в химическом синапсе. Свойства синапсов
Синапс (соединять, смыкать, связывать) – это структурное образование, которое обеспечивает переход возбуждения с нервного волокна на инервируемую клетку.
3. Ионы кальция или его ионизированные комплексы по концентрационному градиенту поступают в нервное окончание (антагонистами кальция являются ионы магния и токсины ботулинуса).
4. Уменьшение электростатических влияний (одноименных зарядов) между пресинаптической мембраной и везикулами.
5. Приближение и слияние везикул с пресинаптической мембраной.
6. Изменение поверхностного натяжения везикул.
8. Выход медиатора в синоптическую щель.
9. Медиатор (возбуждающий в нервно-мышечном синапсе: ацетилхолин) диффундирует через синоптическую щель к рецепторам постсинаптической мембраны.
10. Ацетилхолин вступает во взаимодействие с холинорецепторами (обладают избирательной чувствительностью к ацетилхолину).
11. При одновременном участии ионов кальция и макроэргического фосфата происходят конформационные изменения белковых молекул рецептора.
12. Открываются каналы постсинаптической мембраны для Na или Са.
13. Ионы Na по концентрационному градиенту поступают внутрь воспринимающей возбуждение клетки.
14. Развивается деполяризация –возбуждающий постсинаптический потенциал, который носит местный характер, по форме и свойствам напоминает локальный ответ (не подчиняется закону «все или ничего» и способен суммироваться).
17. Когда он достигает определенной (критической величины) возникают местные токи между возбужденными участками постсинаптической мембраны и невозбужденными участками прилегающей к ней обычной (электровозбудимой) мембраной.
18.На прилегающем участке электровозбудимой мембраны возникает потенциал действия.
Свойства химических синапсов.
1. Нервно-химический механизм передачи возбуждения (передача возбуждения осуществляется с помощью специфического химического вещества – медиатора, который выделяется нервным окончанием и количество которого пропорционально частоте приходящей нервной импульсации).
2. Принцип Дейла (во всех синапсах, образованных нервными окончаниями одного нейрона, выделяется только один вид медиатора – либо возбуждающий, либо тормозный).
3. Одностороннее проведение возбуждения (возбуждение передается только в одном направлении – от пресинаптической мембраны к постсинаптической мембране).
4. Синаптическая задержка (скорость проведения возбуждения в синапсе значительно медленнее, чем в нервном и мышечном волокне).
5. Низкая функциональная лабильность синапса.
6. Трансформация ритма возбуждения (при большой частоте нервных импульсов в пресинаптичоском окончании происходит уменьшение частоты возбуждений в постсинаптических образованиях).
7. Высокая утомляемость (временная потеря работоспособности в результате несоответствия синтеза и расхода медиатора).
Вопросы для самостоятельной подготовки (по учебнику).
1. Медиатор. Виды медиаторов. Свойства медиаторов.
2. Электрические и тормозные синапсы. Особенности передачи сигнала.
3. Пути фармакологической регуляции синаптической передачи возбуждения.